
SNARE complex in axonal guidance and neuroregeneration
2018-04-04 07:40:42FaustoUlloaTizianaCotrufoDeliaRicoloEduardoSorianoSofiaArajo
中國神經(jīng)再生研究(英文版) 2018年3期
Fausto Ulloa, Tiziana Cotrufo, Delia Ricolo, Eduardo Soriano,, So fia J. Araújo,
1 Department of Cell Biology, Physiology and Immunology, School of Biology, and Institute of Neurosciences, University of Barcelona, Barcelona,Spain
2 Centro de Investigación Biomédica en Red sobre Enfermedades Neurodegenerativas (CIBERNED), Instituto de Salud Carlos III (ISCIII), Madrid,Spain
3 Institut de Biologia Molecular de Barcelona (IBMB-CSIC), Parc Cienti fic de Barcelona, Barcelona, Spain
4 Department of Genetics, Microbiology and Statistics, School of Biology, University of Barcelona, Barcelona, Spain
5 Vall d′Hebron Institut de Recerca (VHIR), Barcelona, Spain
Soluble N-Ethylmaleimide Sensitive Factor(NSF) Attachment Protein REceptor (SNARE)Proteins and Neurite Outgrowth
Neuronal differentiation, axonal growth and guidance involve coordinated changes in the cellular cytoskeleton, protein and membrane trafficking processes. Early neurite outgrowth appears to involve protein trafficking machineries responsible for exocytosis to the plasma membrane utilizing many mechanisms that are also found in non-neuronal cells.These membrane trafficking events are usually directed towards several neurites, but can also be switched to concentrate on the growth of a single axon.
Membrane transport to the axonal growth cone is vital for the axon to grow, develop and move. Axonal membrane transport is mediated by the same machinery that governs vesicular trafficking in other parts of the cell. This machinery works in four steps: (1) Budding, in which coat proteins mediate the junction between a membrane donor compartment and motor proteins that direct the transport of vesicles through the cytoskeleton; (2) Movement, when the vesicle moves towards its destination along a cytoskeletal track via molecular motors; (3) Tethering of the vesicle with its target membrane; and (4) Fusion of transmembrane SNAREs (on apposing membranes (Cai et al., 2007).
SNAREs are a large family of small membrane proteins(with more than 60 members in mammalian cells), characterized by the presence, in almost all of them, of the about 60 amino acid SNARE domain. SNARE proteins are related to three different neuronal protein families: vesicle associated membrane protein (VAMPs), Syntaxins, and synaptosomal associated proteins (SNAPs) (Ungar and Hughson,2003). Mechanisms mediated by SNARE proteins are highly conserved and many of the SNARE proteins present in invertebrates are conserved in vertebrates (Teng et al., 2001).SNARE proteins were originally identified for their ability to regulate vesicle release at mature synapses (Chen and Scheller, 2001). In fact, the spontaneous and calcium guided interaction between members of the SNARE family allows for the quantal release of neurotransmitters at the synaptic cleft, which in turn guarantees the efficacy in synaptic transmission (Sudhof, 2013).
Membrane fusion is thought to occur by the formation of a SNARE complex through the association of specific SNARE proteins: a SNARE on a transport vesicle (v-SNARE)assembles with its/their cognate SNARE-binding partner on the appropriate target membrane (t-SNARE). Association between SNAREs is carried out through their SNARE domains and is highly regulated in vivo by several accessory proteins. Members of the Vesicle Associated Membrane Protein (VAMP) subfamily act as v-SNAREs whereas proteins from the Syntaxin and SNAP subfamilies act as t-SNAREs.The best characterized SNARE complex is the one that mediates the Ca2+dependent neurotransmitter exocytosis at the synaptic cleft. It consists of a four-helical bundle formed by the v-SNARE Vamp2, and the t-SNAREs Stx1 and Snap25.However, other types of SNARE complexes can also be formed in different cells or for different functions. For example, spontaneous synaptic vesicle release seems to depend mostly on the v-SNARE Vamp7 instead of Vamp2 (Kavalali,2015). Differentiating the evoked release of neurotransmitters from the spontaneous secretion of vesicles maybe necessary for the communication among neurons. Also, tonic exocytosis, occurring after repetitive stimulation of vesicle release, requires Vamp4 instead of Vamp2 and other types of proteins that allow for the recruiting of the recycling pool of synaptic vesicles and not of the readily releasable pool. In another example, exocytosis in glial cells needs mostly the interaction among Stx1, SNAP23, and VAMP3 (cellubrevin)(Schubert et al., 2011). In addition, diverse SNARE complex composition accounts for the differential sorting of AMPA(at excitatory synapses) and GABA (at inhibitory synapses): [SNAP25-STX1A/B-VAMP2] and [SNAP23-STX1A/B-VAMP2] respectively (Gu et al., 2016). And, regulated exocytosis of the AMPA receptor during long term potentiation involves a unique SNARE fusion machinery containing STX3 (Jurado et al., 2013). So, differential composition/configuration of SNARE complexes can mediate different functions both in the same cell and at distinct cell types (Kasai et al., 2012; van Keimpema and Kroon, 2015).
Neurite outgrowth involves the addition of new membrane, mainly at the tips of elongating axonal processes,coordinated with a dynamic cytoskeletal elongation. Exocytosis of diverse membrane vesicles of around 150 nm in diameter called plasmalemma precursor vesicles (PPVs) or growth cone particles (GCPs) is believed to contribute to is the supply of new membrane in developing axons. In sympathetic neurons and PC12 cells, lysosomal and enlargeosome exocytosis, respectively, have been described to be also involved in neurite outgrowth (Arantes and Andrews, 2006;Colombo et al., 2014). Recently, a non-vesicular mechanism consisting on a lipid flow from the endoplasmatic reticulum(ER) in close apposition to the plasma membrane has been proposed to account for bulk neurite outgrowth in cultured cells (Petkovic et al., 2014). This non-vesicular mechanism involved in membrane expansion requires SNARE proteins Sec22b and Stx1 to generate a SNARE bridge that contributed to plasma membrane expansion (Arantes and Andrews,2006; Petkovic et al., 2014).
Evidence for the implication of SNAREs in neurite outgrowth date from around two decades ago, essentially from in vitro experiments. First, Snap25 requirement in axonal outgrowth was demonstrated by using inhibitory antisense oligonucleotides which prevented neurite elongation (Osen-Sand et al., 1993). Subsequent reports using different approaches, either employing botulinum toxins or protein overexpression confirmed the involvement of Snap25 in neurite outgrowth and sprouting (Morihara et al., 1999;Shirasu et al., 2000). However, mice de ficient for Snap25 did not display neural circuitry defects, suggesting the existence of compensatory mechanisms in action by other members of the SNARE family. Consistent with this idea Stx1a, Stx3,Stx6, Stx13, Snap23, Vamp2, Vamp4 and Vamp7 have also been implicated in neurite outgrowth in roles that vary from membrane trafficking to early endosomal and trans-Golgi network trafficking, exocytosis of endosomes and PPVs (Igarashi et al., 1996; Hirling et al., 2000; Shirasu et al., 2000;Zhou et al., 2000; Martinez-Arca et al., 2001; Darios and Davletov, 2006; Kabayama et al., 2008; Colombo et al., 2014;Grassi et al., 2015).
Evidence for differential SNARE protein expression and function, which can affect the neuronal growth in different cell types, is starting to appear. The best example constitutes the contribution of different isoforms of syntaxin1 protein during embryonic development. In mice, loss of function of STX1A and STX1B give rise to different phenotypes.Whereas STX1A mice have apparently just minor problems,STX1B ones die at early post-natal age. These observations have been attributed to isoform differences, expression pattern and function (Ruiz-Montasell et al., 1996; Kofuji et al.,2014; Mishima et al., 2014; Wu et al., 2015).
Taken together, all these results indicate that more experiments are needed to clarify the involvement of the SNARE complexes in exocytosis during neurite outgrowth.
SNARE Proteins and Endocytosis
As previously mentioned, SNARE proteins have been mainly known to be involved in vesicle release, both in the course of neurotransmission and during the elongation and guidance of the growth cone in development. However, recent findings indicate that SNARE proteins contribute also to the endocytic process, mainly after vesicle release at mature synapses. Endocytosis is needed to recover synaptic vesicles from places on the membrane in which they previously fused. In neurons, two types of endocytosis are generally accepted: a slow, clathrin-dependent endocytosis requiring the classical endocytic proteins as dynamin, AP2 and auxilin(Dittman and Ryan, 2009) and a fast endocytosis, necessary for a faster retrieval of vesicles and proteins as needed for kiss and stay or kiss and run mechanisms of release (Smith et al., 2008). During fast endocytosis synaptic vesicles would keep their identity and also localization. Slow or fast endocytosis are differently activated at synapses matching the activity of the speci fic synapse and its physiological requirements (Smith et al., 2008; Watanabe et al., 2013).
One of the first findings involving SNARE proteins in endocytosis was found in Saccharomyces cerevisiae, where when abolishing the function of a t-SNARE, belonging to the syntaxin protein family, the early steps of the endocytic pathway were imbalanced (Seron et al., 1998). A more critical analysis on the different types of endocytosis in the nervous system showed that fast endocytosis was blocked knocking down Vamp2 in mouse hippocampal synapses(Deak et al., 2004), while abolishing Snap25 at cultured hippocampal neurons did not avoid sucrose induced dye uptake by endocytosis (Bronk et al., 2007). These results appeared to be in con flict with some others in which cleavage of synaptobrevin/Vamp2 with tetanus toxin at a giant nerve terminal, the calyx of Held, blocked slow endocytosis(Hosoi et al., 2009) whilst Snap25 was shown to mainly regulate slow-endocytosis in rat cultured hippocampal synapses(Zhang et al., 2013).
Another study examined all these findings and together with their experiments came to the conclusion that Vamp2 is needed for both slow and fast-endocytosis, as well as Snap25 and Stx1, suggesting the importance of all vesicular and membrane-targeted SNARE in this important process(Xu et al., 2013).
The next steps will be to understand which are the molecular interactions allowing SNARE proteins to differentially participate in exocytosis and endocytosis. At the moment, it has been suggested that the N-terminus of the SNARE motif of Vamp2 binds to the ANTH domain of endocytic adaptors AP180 and Clathrin Assembly Lymphoid Myeloid leukemia(CALM), both of which are involved in endocytosis (Koo et al., 2011; Miller et al., 2011). SNAP-25 is able to bind to intersectin, another endocytic protein, with the same strength it binds to Stx1 (Okamoto et al., 1999). Finally, Stx1 may interact with dynamin, a GTPase mediating vesicle fission after endocytosis (Galas et al., 2000). Overall, more experiments are needed to explore the role of SNARE proteins in endocytosis as it might be of great interest also in elucidating processes occurring during axonal regeneration.
The SNARE Complex in Axonal Guidance in Vertebrates and Invertebrates
Axonal growth cone navigation involves the coordination of cytoskeletal rearrangements with the regulation of adhesion components and membrane trafficking. The growth cone is a unique structure capable of guiding axons to their final destinations. Within the growth cone, extracellular guidance cues are interpreted and then transduced into physical changes and axonal movement. Growth cones are filled with vesicles and express most SNARE and exocyst proteins(Sabo and McAllister, 2003; Yuan et al., 2003; Condeelis et al., 2005; Chernyshova et al., 2011; Fujita et al., 2013). Many reports indicated that vertebrate axon guidance mechanisms require the participation of SNARE-mediated exocytosis for chemoattraction and endocytosis for repulsion (Cotrufo et al., 2011, 2012; Zylbersztejn et al., 2012; Tojima et al.,2014). For example, the vSNARE VAMP2 is required for L1-mediated chemoattraction and for Sema3A-induced chemorepulsion in vivo (Tojima et al., 2007; Zylbersztejn et al.,2012). Compatible with this, Vamp2 de ficient mice show a disorganized corpus callosum similar to the loss of Sema3A(Zylbersztejn et al., 2012). Furthermore, Stx1 and Vamp7 are required for Netrin-1-mediated attraction of axons and migrating neurons, whereas Vamp2 function is dispensable in this process in cultured mouse neurons (Cotrufo et al.,2011, 2012). However, Snap25 and Vamp2 deficient mice show virtually no neural circuitry defects but display a severe alteration of evoked synaptic activity (Schoch et al.,2001; Molnar et al., 2002; Washbourne et al., 2002). And TI-VAMP-de ficient mice display behavioural defects but no alterations in gross brain morphology (Danglot et al., 2012).In addition, the knock-out (KO) mice currently available for Syntaxin-1A (Stx1a) isoform show only mild cognitive defects and a normal brain structure (Fujiwara et al., 2006).And mice KO for the other Stx1 isoform, Stx1b, revealed that STX1B is dispensable for the formation of the mouse neuromuscular junction (NMJ) but required to maintain the efficiency of neurotransmission (Wu et al., 2015). Accordingly, it was suggested that Stx1a and Stx1b are functionally redundant, leading to the need of creation of a double KO mouse by removing both isoforms. Recently, Vardar and colleagues achieved this and showed that Stx1 is essential for the maintenance of developing and mature neurons and also for vesicle docking and neurotransmission (Vardar et al.,2016). However, no analysis of axonal guidance phenotypes was performed in Stx1 double knockout mice. So, currently there is a clear need for more animal models that can clarify the different results obtained from in vivo genetic approaches versus ex vivo petri dish experiments.
Recently, this has been studied using chick and Drosophila melanogaster embryos, two models amenable for gene manipulation followed by in vivo analysis of axonal guidance.In both model systems, the participation of SNARE proteins in the development of the peripheral nervous system (PNS),in particular in the guidance of motor axons has been analysed (Barrecheguren et al., 2017). It was shown that lossof-function of SNARE proteins leads to severe guidance phenotypes in motor axons highlighting a role for neurotransmitter-related SNARE proteins in motor axon guidance in both vertebrates and invertebrates (Barrecheguren et al., 2017).
Drosophila melanogaster presents neural expression of SNARE complex components homologous to the vertebrate complex (DiAntonio et al., 1993; Cerezo et al., 1995; Schulze et al., 1995; Risinger et al., 1997; Moussian et al., 2007).Mutations in components of the core SNARE complex give rise to synaptic transmission and neurotransmitter release phenotypes (Broadie et al., 1995; Schulze et al., 1995; Littleton et al., 1998). In addition, the Drosophila Syntaxin1 homologue, Syntaxin1A (Syx1A) has been reported to affect the properties of neuronal membranes (Schulze and Bellen,1996). Drosophila embryos mutant for Syx1A presented defects in axonal navigation and fasciculation (Barrecheguren et al., 2017). These findings show that defects linked to SNARE protein downregulation are clearly connected to axonal guidance mechanisms. Previous in vitro studies indicated that various SNARE proteins are required for axonal guidance mechanisms linked to Netrin1/DCC and Class III Semaphorins/Plexins (Tojima et al., 2007; Cotrufo et al.,2011, 2012), thereby suggesting that the coupling of guidance receptors to the cell machinery regulating exocytosis is a common mechanism in axonal guidance. Results in fly embryos are in accordance with an interaction of Drosophila Syx1A with frazzled (fra, the fly DCC homolog). However,Syx1A motor axon phenotypes are stronger than the fra phenotypes, and also resemble phenotypes in line with beat-Ia or unc-5 compound guidance mutants (Fambrough and Goodman, 1996; Labrador et al., 2005; Zarin et al., 2014).This suggests that Syx1A may collaborate with axonal guidance pathways other than Netrin/Frazzled. Accordingly,in Drosophila embryos a genetic interaction was detected between Syx1A and Robo pathway components (Barrecheguren et al., 2017).
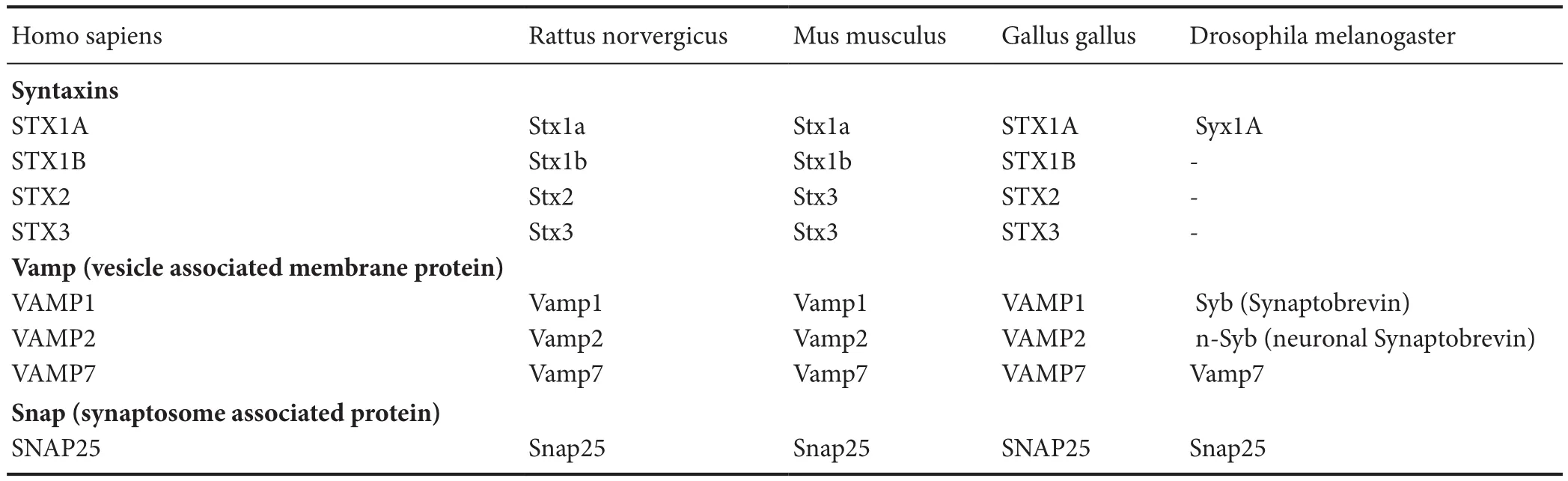
Table 1 GeneBank nomenclature for the Sensitive Factor (NSF) Attachment Protein Receptor (SNARE) complex proteins appearing in this article
Taking together the knowledge generated from using these many different model systems and approaches and taking advantage of the conservation of SNARE protein function between vertebrates and invertebrates, two main models have arisen to incorporate the involvement of SNARE proteins in axonal growth and guidance. These try to incorporate how guidance signals coordinate spatio-temporally the new membrane addition with the cytoskeletal rearrangements despite many of the details not being well understood yet.
One model proposes that SNARE proteins participate in signalling receptor trafficking (Figure 1A). In this model,the blockade of a particular SNARE protein will affect the exocytic delivery and/or the endocytic receptor turnover and, consequently, affect the chemotropic response. This model has been postulated to account for Vamp2 action during Sema3A chemorepulsion (Zylbersztejn et al., 2012)and for the regulation of Robo1 surface expression on commissural axons in vertebrates (Philipp et al., 2012). However, in other systems alternative mechanisms may operate.Another model proposes that SNARE proteins participate in the clustering of receptors onto a speci fic part of the growing growth cone (Figure 1B). This model accounts for the Netrin-1/DCC-mediated attraction of axons depending on Stx1 and Ti-Vamp/Vamp7 (Cotrufo et al., 2011). In this system,the blockade of SNARE proteins does not affect the delivery of the receptor DCC to the plasma membrane. As Stx1 physically associates with DCC and this association is enhanced by the binding of the ligand, Netrin-1 activation of DCC receptors results in ligand dependent clustering of DCC/Stx1 complexes in activated membrane domains. It has been postulated that the membrane expansion at these domains will be produced by the fusion of exocytic vesicles mediated by Stx1—Ti-Vamp/Vamp7 association (Cotrufo et al., 2011).The molecular details explaining why in this scenario Stx1 recruits Ti-Vamp vesicles and not Vamp2 ones, as occurs during neurotransmission, are still not known. There is the possibility that both models can be in play to control axon guidance, the SNARE complex acting in receptor trafficking/turnover but also being responsible for receptor clustering onto speci fic parts of the growing growth cone.
Overall, studies on SNARE proteins in axonal guidance,have presented conflicting data regarding the implication of these proteins directly in axonal guidance at the midline,but seem to suggest that motor axon guidance depends on a functional SNARE complex, which is conserved from invertebrates to vertebrates (Barrecheguren et al., 2017). Current models are missing more studies on the involvement of the SNARE complex in axonal guidance in the midline of both vertebrates and invertebrates.
The SNARE Complex in Neuroregeneration
Injury to the nervous system damages axons, causing their retraction, a widespread loss of synaptic connections, and consequently a deficit of function that can be devastating to the overall organism. Axonal regeneration after damage requires the axon to repair its damaged membrane, redistribute or manufacture what it needs in order to survive, and grow and form new synapses within a more mature, complex environment. For nerve repair to work and regeneration to occur, distinct events have to occur in a coordinate manner. First, anterogradely transported vesicles accumulate at the axons, while others are generated at the severed end to restore a selective barrier to the cut axon. Then, retrograde transport of vesicles along microtubules informs the cell body that damage has occurred in the distal axon.Finally, membrane addition to a newly formed growth cone,or to the axonal membrane is required to promote axonal re-growth and elongation (Tuck and Cavalli, 2010).
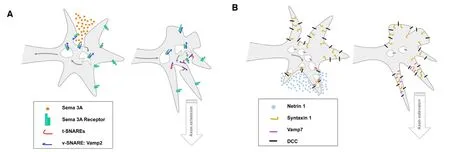
Figure 1 Schematic representation of the different models implicating the Sensitive Factor (NSF) Attachment Protein Receptor (SNARE)complex in axonal guidance.
The molecular mechanisms that support axon repair and growth clearly parallel the mechanisms that mediate synaptic vesicle trafficking and neurotransmitter release within uninjured axons, especially during embryonic development.Specifically, both require calcium, SNARE proteins and their effectors and cytoskeletal remodelling. Members of the SNARE machinery appear to regulate not only vesicle fusion to promote axon resealing but also axonal membrane extension and regrowth (Bloom and Morgan, 2011). For instance,syntaxin13 (Stx13) expression was increased by injury of mouse sciatic nerves in vivo and knockdown of Stx13 in cultured DRG neurons prevented axonal growth and regeneration (Cho et al., 2014).
Most of what we know about SNARE-mediated vesicle exocytosis comes from studies of synapses within uninjured axons (Augustine et al., 1999; Lin and Scheller, 2000; Pang and Sudhof, 2010). Here, calcium entering at the synapse binds to its sensor, synaptotagmin-1. Synaptotagmin then interacts with the SNARE complex, comprising the plasma membrane proteins syntaxin and SNAP-25 and the vesicle-associated membrane protein VAMP-2/ synaptobrevin-2, and in doing so triggers vesicle fusion and neurotransmitter release. Similarly, after injury to squid and cray fish axons, membrane sealing also requires the functions of Syntaxin and Synaptotagmin (Detrait et al., 2000). And regenerating photoreceptors can regulate the expression of a proper set of synaptic vesicle proteins with VAMP being present in all stages of regenerative growth (Yang et al.,2002). Therefore, a model emerges indicating that the critical requirements for ensuring proper membrane sealing and axon extension after injury include iterative bouts of SNARE mediated exocytosis, endocytosis, and functional links between vesicles and the actin cytoskeleton, similar to the mechanisms utilized during synaptic transmission.
Insights into the role of SNAREs in neuroregeneration may come from regenerating model organisms. In geckos(Gekko japonicus), SNAP25 has been shown to be involved in spinal cord regeneration by promoting outgrowth and elongation of neurites (Wang et al., 2012). In tiger salamanders (Ambystoma tigrinum), regenerating adult photoreceptors require autonomous VAMP expression, whereas SNAP-25 is undetectable (Yang et al., 2002). In worms(Caenorhabditis elegans), axonal regeneration is promoted by alternative splicing of Syntaxins (Chen et al., 2016). More studies are needed in these model organisms to bring on a clearer picture of the role of SNAREs in neuroregeneration(Table 1).
Future Directions
Despite the evidence indicating the implication of several SNARE members in neurite outgrowth and axon guidance,their precise role in these processes is far from being well understood. The main challenges for future research in this field are: i) to characterize in detail the precise composition and function of SNARE complexes participating in neurite outgrowth/axon guidance. This could be achieved using imaging techniques such as Fluorescence Resonance Energy Transfer (FRET) or single molecule detection in living cells together with genetic and biochemical strategies; ii) to understand how different SNARE members are selectively regulated and how their action is coordinated with other events relevant in neurite outgrowth/axon guidance; and, iii)to con firm whether and how the nature of cargo molecules,whose transport/sorting is mediated by SNARE complexes,is relevant during the neurite/axon guidance processes. This can be achieved using different model systems, from cell culture to in vivo whole organism analysis. Studies using genetically amenable model organisms such as Drosophila melanogaster in parallel to vertebrate models are starting to prove to be extremely useful in dissecting the function of SNARE proteins in nervous system development (Barrecheguren et al., 2017).
Last but not least, we believe that SNARE function to be extremely relevant in axonal regeneration. Therefore, strategies modulating the activity of SNARE proteins in lesioned axons can potentially be useful in the establishment of new therapies to enhance membrane insertion at the cut axonal edges,thereby potentiating neuronal repair and regeneration.
Author contributions:SJA conceived and wrote the paper. FU, TC and ES wrote the paper and DR constructed figure 1.
Con flicts of interest:None declared.
Financial support:Research in our laboratories was supported by the Ramon y Cajal programme (RYC-2007-00417, RYC-2009-05510) and grants from the Spanish MINECO (SAF2013-42445R and BFU2010-21507) and CIBERNED.
Plagiarism check:Checked twice by iThenticate.
Peer review:Externally peer reviewed.
Open access statement:This is an open access article distributed under the terms of the Creative Commons Attribution-NonCommercial-Shar-eAlike 3.0 License, which allows others to remix, tweak, and build upon the work non-commercially, as long as the author is credited and the new creations are licensed under identical terms.
Open peer review report:
Reviewer:Rong-Kung Tsai, Buddhist Tzu Chi General Hospital, China.
Comments to authors:This review article tries to explain the importance of SNARE complex in axonal guidance and neuroregeneration. It is one of the important issue for neurite regeneration after nerve injury.
Arantes RM, Andrews NW (2006) A role for synaptotagmin VII-regulated exocytosis of lysosomes in neurite outgrowth from primary sympathetic neurons. J Neurosci 26:4630-4637.
Augustine GJ, Burns ME, DeBello WM, Hilfiker S, Morgan JR, Schweizer FE, Tokumaru H, Umayahara K (1999) Proteins involved in synaptic vesicle trafficking. J Physiol 520 Pt 1:33-41.
Barrecheguren PJ, Ros O, Cotrufo T, Kunz B, Soriano E, Ulloa F,Stoeckli ET, Araujo SJ (2017) SNARE proteins play a role in motor axon guidance in vertebrates and invertebrates. Dev Neurobiol 77:963-974.
Bloom OE, Morgan JR (2011) Membrane trafficking events underlying axon repair, growth, and regeneration. Mol Cell Neurosci 48:339-348.
Broadie K, Prokop A, Bellen HJ, O’Kane CJ, Schulze KL, Sweeney ST(1995) Syntaxin and synaptobrevin function downstream of vesicle docking in Drosophila. Neuron 15:663-673.
Bronk P, Deak F, Wilson MC, Liu X, Sudhof TC, Kavalali ET (2007)Differential effects of SNAP-25 deletion on Ca2+-dependent and Ca2+ -independent neurotransmission. J Neurophysiol 98:794-806.
Cai H, Reinisch K, Ferro-Novick S (2007) Coats, tethers, Rabs, and SNAREs work together to mediate the intracellular destination of a transport vesicle. Dev Cell 12:671-682.
Cerezo JR, Jiménez F, Moya F (1995) Characterization and gene cloning of Drosophila syntaxin 1 (Dsynt1): the fruit fly homologue of rat syntaxin 1. Brain Res Mol Brain Res 29:245-252.
Chen L, Liu Z, Zhou B, Wei C, Zhou Y, Rosenfeld MG, Fu XD,Chisholm AD, Jin Y (2016) CELF RNA binding proteins promote axon regeneration in C. elegans and mammals through alternative splicing of Syntaxins. Elife 5:e16072.
Chen YA, Scheller RH (2001) SNARE-mediated membrane fusion. Nat Rev Mol Cell Biol 2:98-106.
Chernyshova Y, Leshchyns’ka I, Hsu SC, Schachner M, Sytnyk V (2011)The neural cell adhesion molecule promotes FGFR-dependent phosphorylation and membrane targeting of the exocyst complex to induce exocytosis in growth cones. J Neurosci 31:3522-3535.
Cho Y, Di Liberto V, Carlin D, Abe N, Li KH, Burlingame AL, Guan S,Michaelevski I, Cavalli V (2014) Syntaxin13 expression is regulated by mammalian target of rapamycin (mTOR) in injured neurons to promote axon regeneration. J Biol Chem 289:15820-15832.
Colombo F, Racchetti G, Meldolesi J (2014) Neurite outgrowth induced by NGF or L1CAM via activation of the TrkA receptor is sustained also by the exocytosis of enlargeosomes. Proc Natl Acad Sci U S A 111:16943-16948.
Condeelis J, Singer RH, Segall JE (2005) The great escape: when cancer cells hijack the genes for chemotaxis and motility. Annu Rev Cell Dev Biol 21:695-718.
Cotrufo T, Andrés RM, Ros O, Pérez-Brangulí F, Muhaisen A, Fuschini G, Martínez R, Pascual M, Comella JX, Soriano E (2012) Syntaxin 1 is required for DCC/Netrin-1-dependent chemoattraction of migrating neurons from the lower rhombic lip. Eur J Neurosci 36:3152-3164.
Cotrufo T, Perez-Branguli F, Muhaisen A, Ros O, Andres R, Baeriswyl T,Fuschini G, Tarrago T, Pascual M, Urena J, Blasi J, Giralt E, Stoeckli ET, Soriano E (2011) A signaling mechanism coupling netrin-1/deleted in colorectal cancer chemoattraction to SNARE-mediated exocytosis in axonal growth cones. J Neurosci 31:14463-14480.
Danglot L, Zylbersztejn K, Petkovic M, Gauberti M, Meziane H, Combe R, Champy MF, Birling MC, Pavlovic G, Bizot JC, Trovero F, Della Ragione F, Proux-Gillardeaux V, Sorg T, Vivien D, D’Esposito M,Galli T (2012) Absence of TI-VAMP/Vamp7 leads to increased anxiety in mice. J Neurosci 32:1962-1968.
Darios F, Davletov B (2006) Omega-3 and omega-6 fatty acids stimulate cell membrane expansion by acting on syntaxin 3. Nature 440:813-817.
Deak F, Schoch S, Liu X, Sudhof TC, Kavalali ET (2004) Synaptobrevin is essential for fast synaptic-vesicle endocytosis. Nat Cell Biol 6:1102-1108.
Detrait E, Eddleman CS, Yoo S, Fukuda M, Nguyen MP, Bittner GD,Fishman HM (2000) Axolemmal repair requires proteins that mediate synaptic vesicle fusion. J Neurobiol 44:382-391.
DiAntonio A, Burgess RW, Chin AC, Deitcher DL, Scheller RH,Schwarz TL (1993) Identi fication and characterization of Drosophila genes for synaptic vesicle proteins. J Neurosci 13:4924-4935.
Dittman J, Ryan TA (2009) Molecular circuitry of endocytosis at nerve terminals. Annu Rev Cell Dev Biol 25:133-160.
Fambrough D, Goodman CS (1996) The Drosophila beaten path gene encodes a novel secreted protein that regulates defasciculation at motor axon choice points. Cell 87:1049-1058.
Fujita A, Koinuma S, Yasuda S, Nagai H, Kamiguchi H, Wada N,Nakamura T (2013) GTP hydrolysis of TC10 promotes neurite outgrowth through exocytic fusion of Rab11- and L1-containing vesicles by releasing exocyst component Exo70. PLoS One 8:e79689.
Fujiwara T, Mishima T, Kofuji T, Chiba T, Tanaka K, Yamamoto A,Akagawa K (2006) Analysis of knock-out mice to determine the role of HPC-1/syntaxin 1A in expressing synaptic plasticity. J Neurosci 26:5767-5776.
Galas MC, Chasserot-Golaz S, Dirrig-Grosch S, Bader MF (2000)Presence of dynamin--syntaxin complexes associated with secretory granules in adrenal chromaffin cells. J Neurochem 75:1511-1519.
Grassi D, Plonka FB, Oksdath M, Guil AN, Sosa LJ, Quiroga S (2015)Selected SNARE proteins are essential for the polarized membrane insertion of igf-1 receptor and the regulation of initial axonal outgrowth in neurons. Cell Discov 1:15023.
Gu Y, Chiu SL, Liu B, Wu PH, Delannoy M, Lin DT, Wirtz D, Huganir RL (2016) Differential vesicular sorting of AMPA and GABAA receptors. Proc Natl Acad Sci U S A 113:E922-931.
Hirling H, Steiner P, Chaperon C, Marsault R, Regazzi R, Catsicas S(2000) Syntaxin 13 is a developmentally regulated SNARE involved in neurite outgrowth and endosomal trafficking. Eur J Neurosci 12:1913-1923.
Hosoi N, Holt M, Sakaba T (2009) Calcium dependence of exo- and endocytotic coupling at a glutamatergic synapse. Neuron 63:216-229.
Igarashi M, Kozaki S, Terakawa S, Kawano S, Ide C, Komiya Y (1996)Growth cone collapse and inhibition of neurite growth by Botulinum neurotoxin C1: a t-SNARE is involved in axonal growth. J Cell Biol 134:205-215.
Jurado S, Goswami D, Zhang Y, Molina AJ, Sudhof TC, Malenka RC(2013) LTP requires a unique postsynaptic SNARE fusion machinery. Neuron 77:542-558.
Kabayama H, Tokushige N, Takeuchi M, Mikoshiba K (2008) Syntaxin 6 regulates nerve growth factor-dependent neurite outgrowth. Neurosci Lett 436:340-344.
Kasai H, Takahashi N, Tokumaru H (2012) Distinct initial SNARE configurations underlying the diversity of exocytosis. Physiol Rev 92:1915-1964.
Kavalali ET (2015) The mechanisms and functions of spontaneous neurotransmitter release. Nat Rev Neurosci 16:5-16.
Kofuji T, Fujiwara T, Sanada M, Mishima T, Akagawa K (2014) HPC-1/syntaxin 1A and syntaxin 1B play distinct roles in neuronal survival. J Neurochem 130:514-525.
Koo SJ, Markovic S, Puchkov D, Mahrenholz CC, Beceren-Braun F,Maritzen T, Dernedde J, Volkmer R, Oschkinat H, Haucke V (2011)SNARE motif-mediated sorting of synaptobrevin by the endocytic adaptors clathrin assembly lymphoid myeloid leukemia (CALM)and AP180 at synapses. Proc Natl Acad Sci U S A 108:13540-13545.
Labrador JP, O’keefe D, Yoshikawa S, McKinnon RD, Thomas JB,Bashaw GJ (2005) The homeobox transcription factor even-skipped regulates netrin-receptor expression to control dorsal motor-axon projections in Drosophila. Curr Biol 15:1413-1419.
Lin RC, Scheller RH (2000) Mechanisms of synaptic vesicle exocytosis.Annu Rev Cell Dev Biol 16:19-49.
Littleton JT, Chapman ER, Kreber R, Garment MB, Carlson SD,Ganetzky B (1998) Temperature-sensitive paralytic mutations demonstrate that synaptic exocytosis requires SNARE complex assembly and disassembly. Neuron 21:401-413.
Martinez-Arca S, Coco S, Mainguy G, Schenk U, Alberts P, Bouille P,Mezzina M, Prochiantz A, Matteoli M, Louvard D, Galli T (2001) A common exocytotic mechanism mediates axonal and dendritic outgrowth. J Neurosci 21:3830-3838.
Miller SE, Sahlender DA, Graham SC, Honing S, Robinson MS, Peden AA, Owen DJ (2011) The molecular basis for the endocytosis of small R-SNAREs by the clathrin adaptor CALM. Cell 147:1118-1131.
Mishima T, Fujiwara T, Sanada M, Kofuji T, Kanai-Azuma M, Akagawa K (2014) Syntaxin 1B, but not syntaxin 1A, is necessary for the regulation of synaptic vesicle exocytosis and of the readily releasable pool at central synapses. PLoS One 9:e90004.
Molnar Z, Lopez-Bendito G, Small J, Partridge LD, Blakemore C, Wilson MC (2002) Normal development of embryonic thalamocortical connectivity in the absence of evoked synaptic activity. J Neurosci 22:10313-10323.
Morihara T, Mizoguchi A, Takahashi M, Kozaki S, Tsujihara T, Kawano S, Shirasu M, Ohmukai T, Kitada M, Kimura K, Okajima S, Tamai K, Hirasawa Y, Ide C (1999) Distribution of synaptosomal-associated protein 25 in nerve growth cones and reduction of neurite outgrowth by botulinum neurotoxin A without altering growth cone morphology in dorsal root ganglion neurons and PC-12 cells. Neuroscience 91:695-706.
Moussian B, Veerkamp J, Müller U, Schwarz H (2007) Assembly of the Drosophila larval exoskeleton requires controlled secretion and shaping of the apical plasma membrane. Matrix Biol 26:337-347.
Okamoto M, Schoch S, Sudhof TC (1999) EHSH1/intersectin, a protein that contains EH and SH3 domains and binds to dynamin and SNAP-25. A protein connection between exocytosis and endocytosis? J Biol Chem 274:18446-18454.
Osen-Sand A, Catsicas M, Staple JK, Jones KA, Ayala G, Knowles J,Grenningloh G, Catsicas S (1993) Inhibition of axonal growth by SNAP-25 antisense oligonucleotides in vitro and in vivo. Nature 364:445-448.
Pang ZP, Sudhof TC (2010) Cell biology of Ca2+-triggered exocytosis.Curr Opin Cell Biol 22:496-505.
Petkovic M, Jemaiel A, Daste F, Specht CG, Izeddin I, Vorkel D, Verbavatz JM, Darzacq X, Triller A, Pfenninger KH, Tareste D, Jackson CL, Galli T (2014) The SNARE Sec22b has a non-fusogenic function in plasma membrane expansion. Nat Cell Biol 16:434-444.
Philipp M, Niederko fler V, Debrunner M, Alther T, Kunz B, Stoeckli ET (2012) RabGDI controls axonal midline crossing by regulating Robo1 surface expression. Neural Dev 7:36.
Risinger C, Deitcher DL, Lundell I, Schwarz TL, Larhammar D (1997)Complex gene organization of synaptic protein SNAP-25 in Drosophila melanogaster. Gene 194:169-177.
Ruiz-Montasell B, Aguado F, Majo G, Chapman ER, Canals JM, Marsal J, Blasi J (1996) Differential distribution of syntaxin isoforms 1A and 1B in the rat central nervous system. Eur J Neurosci 8:2544-2552.
Sabo SL, McAllister AK (2003) Mobility and cycling of synaptic protein-containing vesicles in axonal growth cone filopodia. Nat Neurosci 6:1264-1269.
Schoch S, Deak F, Konigstorfer A, Mozhayeva M, Sara Y, Sudhof TC,Kavalali ET (2001) SNARE function analyzed in synaptobrevin/VAMP knockout mice. Science 294:1117-1122.
Schubert V, Bouvier D, Volterra A (2011) SNARE protein expression in synaptic terminals and astrocytes in the adult hippocampus: a comparative analysis. Glia 59:1472-1488.
Schulze KL, Bellen HJ (1996) Drosophila syntaxin is required for cell viability and may function in membrane formation and stabilization.Genetics 144:1713-1724.
Schulze KL, Broadie K, Perin MS, Bellen HJ (1995) Genetic and electrophysiological studies of Drosophila syntaxin-1A demonstrate its role in nonneuronal secretion and neurotransmission. Cell 80:311-320.
Seron K, Tieaho V, Prescianotto-Baschong C, Aust T, Blondel MO,Guillaud P, Devilliers G, Rossanese OW, Glick BS, Riezman H, Keranen S, Haguenauer-Tsapis R (1998) A yeast t-SNARE involved in endocytosis. Mol Biol Cell 9:2873-2889.
Shirasu M, Kimura K, Kataoka M, Takahashi M, Okajima S, Kawaguchi S, Hirasawa Y, Ide C, Mizoguchi A (2000) VAMP-2 promotes neurite elongation and SNAP-25A increases neurite sprouting in PC12 cells. Neurosci Res 37:265-275.
Smith SM, Renden R, von Gersdorff H (2008) Synaptic vesicle endocytosis: fast and slow modes of membrane retrieval. Trends Neurosci 31:559-568.
Sudhof TC (2013) A molecular machine for neurotransmitter release:synaptotagmin and beyond. Nat Med 19:1227-1231.
Teng FY, Wang Y, Tang BL (2001) The syntaxins. Genome Biol 2:REVIEWS3012.
Tojima T, Itofusa R, Kamiguchi H (2014) Steering neuronal growth cones by shifting the imbalance between exocytosis and endocytosis.J Neurosci 21:7165-7178.
Tojima T, Akiyama H, Itofusa R, Li Y, Katayama H, Miyawaki A, Kamiguchi H (2007) Attractive axon guidance involves asymmetric membrane transport and exocytosis in the growth cone. Nat Neurosci 10:58-66.
Tuck E, Cavalli V (2010) Roles of membrane trafficking in nerve repair and regeneration. Commun Integr Biol 3:209-214.
Ungar D, Hughson FM (2003) SNARE protein structure and function.Annu Rev Cell Dev Biol 19:493-517.
van Keimpema L, Kroon T (2015) Do SNARE protein isoforms determine fusion pore characteristics? J Neurosci 35:11459-11461.
Vardar G, Chang S, Arancillo M, Wu YJ, Trimbuch T, Rosenmund C(2016) Distinct functions of syntaxin-1 in neuronal maintenance,synaptic vesicle docking, and fusion in mouse neurons. J Neurosci 36:7911-7924.
Wang Y, Dong Y, Song H, Liu Y, Liu M, Yuan Y, Ding F, Gu X, Wang Y (2012) Involvement of gecko SNAP25b in spinal cord regeneration by promoting outgrowth and elongation of neurites. Int J Biochem Cell Biol 44:2288-2298.
Washbourne P, Thompson PM, Carta M, Costa ET, Mathews JR, Lopez-Bendito G, Molnar Z, Becher MW, Valenzuela CF, Partridge LD, Wilson MC (2002) Genetic ablation of the t-SNARE SNAP-25 distinguishes mechanisms of neuroexocytosis. Nat Neurosci 5:19-26.Watanabe S, Rost BR, Camacho-Perez M, Davis MW, Sohl-Kielczynski B, Rosenmund C, Jorgensen EM (2013) Ultrafast endocytosis at mouse hippocampal synapses. Nature 504:242-247.
Wu Y-J, Tejero R, Arancillo M, Vardar G, Korotkova T, Kintscher M, Schmitz D, Ponomarenko A, Tabares L, Rosenmund C (2015)Syntaxin 1B is important for mouse postnatal survival and proper synaptic function at the mouse neuromuscular junctions. J Neurophysiol 114:2404-2417.
Xu J, Luo F, Zhang Z, Xue L, Wu XS, Chiang HC, Shin W, Wu LG(2013) SNARE proteins synaptobrevin, SNAP-25, and syntaxin are involved in rapid and slow endocytosis at synapses. Cell Rep 3:1414-1421.
Yang H, Standifer KM, Sherry DM (2002) Synaptic protein expression by regenerating adult photoreceptors. J Comp Neurol 443:275-288.
Yuan XB, Jin M, Xu X, Song YQ, Wu CP, Poo MM, Duan S (2003) Signalling and crosstalk of Rho GTPases in mediating axon guidance.Nat Cell Biol 5:38-45.
Zarin AA, Asadzadeh J, Labrador JP (2014) Transcriptional regulation of guidance at the midline and in motor circuits. Cell Mol Life Sci 71:419-432.
Zhang Z, Wang D, Sun T, Xu J, Chiang HC, Shin W, Wu LG (2013)The SNARE proteins SNAP25 and synaptobrevin are involved in endocytosis at hippocampal synapses. J Neurosci 33:9169-9175.
Zhou Q, Xiao J, Liu Y (2000) Participation of syntaxin 1A in membrane trafficking involving neurite elongation and membrane expansion. J Neurosci Res 61:321-328.
Zylbersztejn K, Petkovic M, Burgo A, Deck M, Garel S, Marcos S,Bloch-Gallego E, Nothias F, Serini G, Bagnard D, Binz T, Galli T(2012) The vesicular SNARE Synaptobrevin is required for Semaphorin 3A axonal repulsion. J Cell Biol 196:37-46.

- 中國神經(jīng)再生研究(英文版)的其它文章
- The biological clock: future of neurological disorders therapy
- Optic radiation injury in patients with aneurismal subarachnoid hemorrhage: a preliminary diffusion tensor imaging report
- Regulatory role of calpain in neuronal death
- The ROCK pathway inhibitor Y-27632 mitigates hypoxia and oxidative stress-induced injury to retinal Müller cells
- Combined acupuncture and HuangDiSan treatment affects behavior and synaptophysin levels in the hippocampus of senescence-accelerated mouse prone 8 after neural stem cell transplantation
- Endoplasmic reticulum stress transducer old astrocyte speci fically induced substance contributes to astrogliosis after spinal cord injury