
The Differing Fortunes and miRNA Clusters Between Human Astrocytes and Neurons After Endoplasmic Reticulum Stress With Downregulation of EIF2B5*
2022-11-22 10:44:06CHENNaWANGJingMinJIANGYuWuWUYe
生物化學(xué)與生物物理進(jìn)展 2022年11期
CHEN Na,WANG Jing-Min,JIANG Yu-Wu,WU Ye**
(1)Department of Pediatrics,Peking University First Hospital,Beijing 100034,China;2)Department of Pediatrics,Beijing Tiantan Hospital,Capital Medical University,Beijing 100070,China)
Abstract Objective Human astrocytes and human neurons were transfected with EIF2B5-RNAi vectors or blank as cell model to investigate the causes that white matter glial cells selectively involved in vanishing white matter disease.Methods Apoptosis and cell viability were measured in human astrocytes and neurons with EIF2B5-RNAi or blank vector before or after the stimulation of endoplasmic reticulum stress(ERS).The sequence of known and unknown microRNAs was performed to screen those participated in regulating the response of ERS.Results Under the baseline condition,apoptosis rates were detected higher in human astrocytes with EIF2B5-RNAi and cell viability decreased significantly than the wild-type and neurons.More pieces of miRNAs regulated the ERS rescue in astrocytes with blank vectors than the EIF2B5-RNAi group.In the cluster analysis,5 pieces of miRNAs were identified as key components in pathway network.Conclusion The recovery of human astrocytes from ERS may be more dependent on the numerous miRNAs than neurons for cell proliferation and differentiation.The eroded clusters of miRNAs may promote spontaneous apoptosis and cell viability decrease in human astrocytes with EIF2B5-RNAi.As a result,it reduced the chance of survival in human astrocytes with EIF2B5-RNAi after ERS crisis.
Key words vanishing white matter disease,human astrocytes,human neurons,endoplasmic reticulum stress,apoptosis,miRNAs differences
Vanishing white matter disease(VWM)is an autosomal recessive leukodystrophy that develop all ages and life threatening at an early onset age.It is caused by mutations in any of the genes ofEIF2B1-5encoding the 5 subunits of eukaryotic translation initiation factor 2B(eIF2B).EIF2B1-5are housekeeping genes and expressed ubiquitously,however,VWM manifests as a leukoencephalopathy with progressive cerebral white matter rarefaction and rapid neurological deterioration as the classical phenotype.VWM patients are sensitive to stress conditions as febrile infections,mild head trauma and acute fright,manifested as chronic progressive motor deterioration and relatively mild mental decline[1-2].Pathological findings from VWM patients include thin myelin sheaths and gelatinous,scarce white matter in the focal zone but the grey matter is generally spared or greatly preserved[2-7].On macroscopic examination showed foamy oligodendrocytes, meager astrogliosis with dysmorphic astrocytes and loss of oligodendrocytes by apoptosis.Mutations impair the activity of eIF2B and its function to couple protein synthesis to the cellular demands,leading to sustained activation of the unfolded protein response(UPR),especially during stress conditions[6,8].In our previous study,eIF2B mutant oligodendrocytes are susceptible to endoplasmic reticulum stress(ERS)stimulation.The UPR pathway is activated under basic condition and overactivated after ERS,with the depression of autophagy,resulting to the inability to terminate ERS and a large amount of apoptotic death in oligodendrocytes[9-10].MicroRNAs(miRNA)play important parts in cell proliferation and differentiation for promoting cell survival.Several chemicals have been identified work through miRNAs[11-13].However,up to now limited studies on the miRNAs in the pathogenesis of VWM[11-13].Astrocytes are essential in central nervous system(CNS)for the paucity of astrogliosis and abnormal morphology of astrocytes in VWM patient leading to the vanished white matter[2].In this study,human astrocytes and human neurons were transfected withEIF2B5-RNAi or blank vector for comparison under basic condition or after ERS.To get a better understanding of the reasons that glial selectively involved in VWM,different destinies were measured between neurons and astrocytes withEIF2B5knockdown after ERS stimulation and the clusters of upstream miRNAs were also detected.
1 Materials and methods
1.1 Culture of primary human astrocytes and neurons
Primary human astrocytes(Catalog# 1800)(HA)and primary human neurons(Catalog# 1520-10),and the culture medium for human astrocytes(Catalog#1801)and human neurons(Catalog# 1521)were purchased from ScienCell Research Laboratories,America.Cell culture and passage were the same as those of normal adherent cell culture.First the cell culture plates were covered with 0.1 g/L polylysine(PLL)(Sigma),37°C 30 min,washing the culture plate twice with sterile ddH2O,removing the residual liquid thoroughly,plating with cells for spare,changing medium after cell adhering to the plates after 16-18 h.
1.2 Vectors of EIF2B5-RNAi and lentiviral transduction
1.2.1Vectors ofEIF2B5-RNAi
TheEIF2B5lentiviral vector sequence(NM_003907)was referred from the overexpression ofEIF2B5-targeted RNAi(pBSU6-581i),published by J?rg Dietrich for human glial precursor cells(hGPC)[5].The carrier is pGLV2-U6-Puro constructed by Shanghai GenePharma Co.,Ltd.TheEIF2B5mRNA was degraded when the vector of pGLV2-U6 581i RNAi transfected in cells,calledEIF2B5silence.TheEIF2B5-targeted RNAi(pBSU6-581i)is as follows.EIF2B5hs581i forward:5'-GATCCCCCTCGTTGCCACGAAGACAATTCAAGAGATTGTCTTCGT GGCAACGAGTTTTTGGAAA-3';EIF2B5hs581i reverse:5'-AGCTTTTCCAAAAACTCGTTGCCACGAAGACAATCTCTTGAATTGTCTTCGTGGCAA CGAGGG-3'.
1.2.2Transduction efficiency ofEIF2B5-RNAi
Transfection efficiency was validated with realtime PCR forEIF2B5transcription and Western blot for FLAG-tag.The vectors withEIF2B5-RNAi or blank were transfected into human astrocytes with an optimized condition.72 h after transfection,the cells were harvested for the following studies.The forward primer ofEIF2B5:5'-TGGAGTGGAGGTTCGATATGAT-3';the reverse primer:5'-GGTTCCCTAGGATCTCCTCAT-3'.
1.3 miRNA sequencing
Eight groups of lentiviral vectors were transferred human astrocytes or human neurons respectively.The miRNAs were compared pairwise before and after ERS.1 μmol/L Thapsigargin(TG,#T9033,Sigma-Aldrich,USA)was used for the stimulation of ERS.According to the screening strategy,differences greater than 2 times were considered significant.The marks T01 to T04 represent human astrocytes(T01 and T02 with TG,T03 and T04 without TG);T05-T08 are neuron groups(T05 and T07 are TG group,T06 and T08 without TG).Vector(v)representing blank vector andEIF2B5-RNAi(i)representsEIF2B5-RNAi groups for neuron or astrocyte.The marks include T01(Astrocyte vTG),T02(Astrocyte iTG),T03(Astrocyte i),T04(Astrocyte v),T05(Neuron iTG),T06(Neuron i),T07(Neuron vTG)and T08(Neuron v).
High-throughput was performed by illumina HiSeq/MiSeq(Beijing Berry Genomics Co.,Ltd).The original image date files obtained from illumine sequencing was analyzed by Base Calling and transferred to Sequenced Reads(raw reads).They were documented as the file format of FASTQ(fq),which includes reads and sequencing quality information.The primary analysis of raw data was completed by the procedure of de-coupling,de-low quality,de-contamination and statistical sequence length distribution.Clean reads obtained from primary analysis were compared with the ncRNA library,which were classified and annotated,removing rNRA,tRNA,snRNA,snoRNA.The reads were then aligned to the reference genome and miRNA base,calculating miRNA expression,annotating the known miRNAs and predicting new miRNAs for further analysis.
The length of clean reads distribution:the number of 18-31 nt clean reads and respective percentage of total were calculated,plotting the length distribution of reads.Generally,the length of small RNA ranges 18-31 nt,and the peak of distribution helps us to judge the type of small RNAs.For example,miRNA was concentrated at 21 or 22 nt,siRNA at 24 nt and piRNA at 30 nt.The Venn diagram method was used to visually display the differences in the number of known miRNAs,predicting new miRNAs in multiple samples,also with RFAM alignment,base bias analysis and clean reads after removing rRNA,scRNA,snRNA,snoRNA,tRNA and ncRNA compared from the reference genome.The total matched reads were compared with the corresponding genome to predict the known miRNA target genes.
1.4 Detection of cell apoptosis and viability for the tolerance of ERS
Rates of apoptosis and cell viability were measured by Annexin V-FITC and quantitated by flow cytometry(FCM).FCM detects the rates of cell apoptosis under basic condition or after ERS stimulation.Astrocytes or neurons were doublelabeled with Annexin V-FITC/propidium iodide(PI)by apoptosis detection kit(#KGA108,KeyGEN BioTECH, Beijing, China). Normally, the phosphatidylserine(PS)is only distributed in the inner side of the lipid bilayer of the cell membrane,in the early stage of apoptosis however,PS turns from the inner side of the lipid membrane to the outside.Annexin V has a high affinity for phosphatidyl serine,so it could bind to the membrane of early apoptotic cells through the PS exposed on the outside of the cell.PI is a nuclei acid dye which could not pass through the intact cell membrane but the late apoptotic cells and dead cells could be dyed red.Therefor the use of Annexin V and PI helps to distinguish cells in different status of apoptosis from dead cells,as apoptotic cells(Annexin V+,PI-),necrotic cells(Annexin V+,PI+),no apoptotic or necrotic cells(Annexin V-,PI-).Astrocytes or neurons without transfection were set as baseline(0 h),comparing with theEIF2B5-RNAi or blank vector transfected cells.Cell viability was measured under basic condition and at different time points of 8 h,24 h and 48 h after ERS stimulation with the Cell Counting Kit-8(CCK-8)(#CK04,Dojindo Molecular Technologies,Japan).Statistical differences between two groups were determined by two-tailed unpairedt-test.One-way analysis of variance was for more than two groups.P<0.05 was considered significantly different(*P<0.05,**P<0.01,***P<0.001).The results were repeated 3 times separately.
2 Resul ts
2.1 The primary human astrocytes with EIF2B5 knockdown showed increased apoptosis rates and decreased cell viability under basic condition and further reduced the cell survival after ERS stimulation
Primary human astrocytes transfected with PGLV2-U6 581i RNA interference vector ofEIF2B5were compared with astrocytes with PGLV2-U6 control vector.The changes of cell viability and apoptosis level were observed at basal state(natural culture)and ERS stimulation(1 μmol/L TG).In basal state,human astrocytes without transfection and transfected with control vector began to proliferate at 48-72 h with the extension of culture time,and the cell viability increased.However,the viability of astrocytes withEIF2B5knockdown began to decline at 8 h of natural culture,and remained at a low level at 24,48 and 72 h(Figure 1a).Astrocytes without transfection or ERS stimulation(TG-)were modulated as the baseline of apoptosis.FCM results showed that the apoptosis level of primary human astrocytes transfected withEIF2B5PGLV2-U6 581i RNA interference vector was significantly higher than that of PGLV2-U6 control vector group after 48 h of natural culture(Figure 1c).
After ERS stimulation,the viability of astrocytes decreased to some extent in the non-transfected group,the control vector transfected group and theEIF2B5knockdown group.The cell viability decreased rapidly in theEIF2B5knockdown group at 8-24 h after ERS stimulation,and remained at a low level at 48-72 h after ERS stimulation(Figure 1b).The apoptotic rates ofEIF2B5knockdown group were significantly higher than control vector group after ERS induction(Figure 1c).
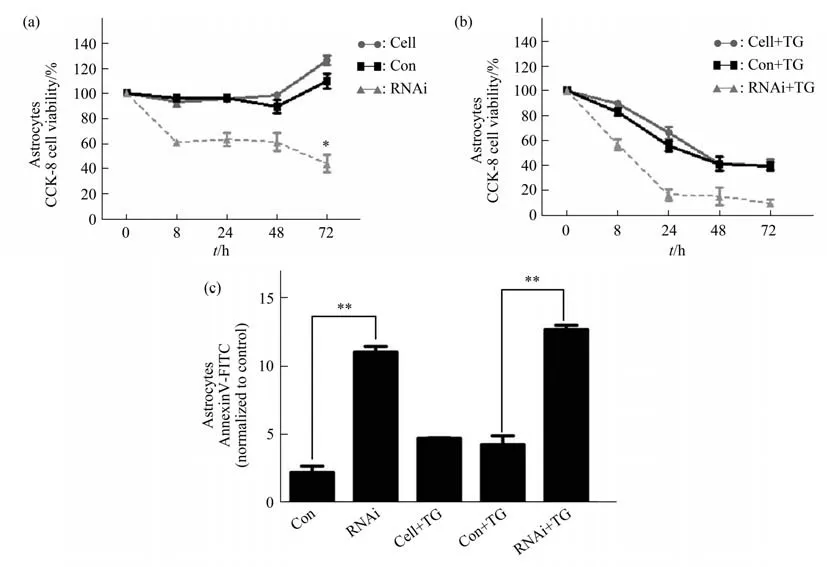
Fig.1 Detection of viability and apoptosis rate in primary human astrocytes under basal condition and after ERS
2.2 Human neurons with EIF2B5 knockdown did not show increased apoptosis and decreased viability with culture time in the basal condition
Cell viability is not reduced in neurons transfected withEIF2B5pGLV2-U6 581i RNA or pGLV2-U6 blank vector groups in basal condition(Figure 2a).FCM analysis showed that there was no significant difference in apoptosis level betweenEIF2B5knockdown group and control vector group after 48 h of natural culture(Figure 2c).
After 8 h of ERS stimulation,the cell viability ofEIF2B5knockdown group,control vector group and not transfected group decreased rapidly compared with the basal state.There was no significant difference in cell viability between theEIF2B5knockdown group and the control vector group,or between the control vector group and the group without transfection(Figure 2b).FCM showed thatEIF2B5knockdown neurons showed a significant increase in apoptosis after ERS stimulation(Figure 2c).
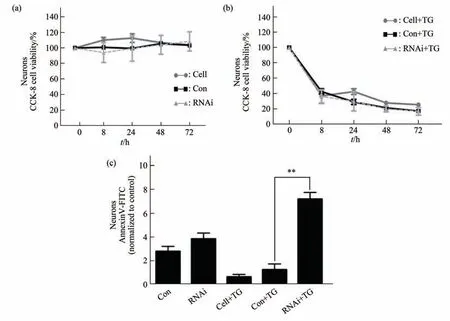
Fig.2 Cell viability and apoptosis level of human neurons under basic condition and after ERS
2.3 Astrocytes require more miRNAs to participate in regulation than neurons in the basal condition and the number of miRNAs involved in regulation decreased significantly after EIF2B5-RNAi
More pieces of known and unknown miRNAs were measured participated in the regulation after ERS stimulation in human astrocytes,but the number of miRNAs involved reduced after the transfection ofEIF2B5-RNAi vectors.Among the known miRNA data,there were 345 differentially expressed miRNAs between wild-type andEIF2B5-RNAi transfected human astrocytes in the basal state.
It was detected 350 miRNAs in wild-type human astrocytes after ERS stimulation,while only 5 differential miRNAs inEIF2B5-RNAi human astrocytes.In human neurons,there were 43 different miRNAs between wild-type andEIF2B5-knock down human neurons in the basal state.After ERS stimulation,46 differentially expressed miRNAs were obtained in wild-type human neurons and human neurons withEIF2B5-knock detected 15 differentially expressed miRNAs.
The miRNAs that engaged in the regulation after ERS crisis were screening in astrocytes and neurons transfected withEIF2B5-RNAi(Figure 3a).N1,the miRNA difference present in wild-type andEIF2B5-RNAi astrocytes;N2,miRNA changes after ERS stimulation in wild-type astrocytes;N3,miRNAs involved in ERS regulation ofEIF2B5-RNAi transfected astrocytes;N2 was compared with N3 to obtain N4,and then the differences between wild-type and RNAi human astrocytes before ERS stimulation were compared to obtain N9(miRNAs differences in astrocytes with or withoutEIF2B5-RNAi transfection before and after ERS).Parallel experiments to compare human neurons:N5 is the miRNAs difference between wild-type andEIF2B5-RNAi human neurons.N6 and N7 respectively represent the difference in miRNA expression between wild-type andEIF2B5-RNAi human neurons after ERS stimulation.After N8,N10 was obtained by comparing the difference between wild-type andEIF2B5-RNAi human neurons before ERS stimulation(miRNA differences in human neurons with wild type orEIF2B5-RNAi vectors after ERS).N9 and N10 were compared to determine which miRNAs were significantly changed in N11,which is the final miRNA changes in the wild-type andEIF2B5-RNAi human astrocytes and human neurons after ERS stimulation.Then go back to the original database and find out which procedure the changes came from(Figure 3b).
The results of N11 screened above were substituted in to find common and unique miRNAs among the different samples(Figure 3b).Finding out the natural differences in astrocytes and neurons,N12 was harvested.Comparing the miRNA differences between wild-type astrocytes and neurons,finding out the same miRNAs we obtained in N11,getting 58 pieces of miRNA as N12 for background data.N13 was obtained after screening out the miRNAs differences between human astrocytes and human neurons transfected withEIF2B5-RNAi,finding out the same miRNAs in N11.N13 is derived from the miRNA differences between human astrocytes and human neurons caused byEIF2B5-RNAi factor,and the number of 190 pieces of miRNA as N14 is obtained by subtracting the natural difference between the two types of cells.Identifying the different pieces of miRNA expression between human astrocytes and neurons withEIF2B5-RNAi after ERS stimulation,and then to find the same miRNA in N11,getting N15,removing the N12 part,obtaining the number of 196 pieces of miRNAs as N16.
The selected miRNAs were subjected to signal pathway cluster analysis and finally 5 known miRNAs,hsa-miR-335-5p,hsa-miR-92a-3p,hsa-miR-548aj-5p,hsa-miR-548g-5p,hsa-miR-548x-5p,were emerged from the miRNA data pool,which play key roles in the connection of multiple signal pathways,participating in cell proliferation and differentiation(Figure 4).
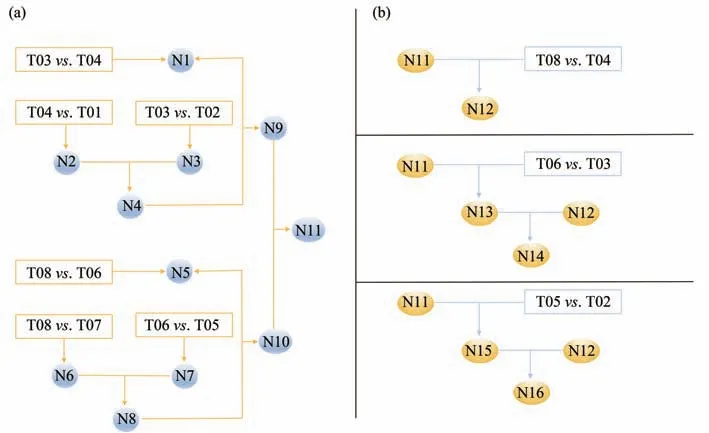
Fig.3 The miRNA comparison strategies of astrocytes and neurons in basal state and after ERS in different groups
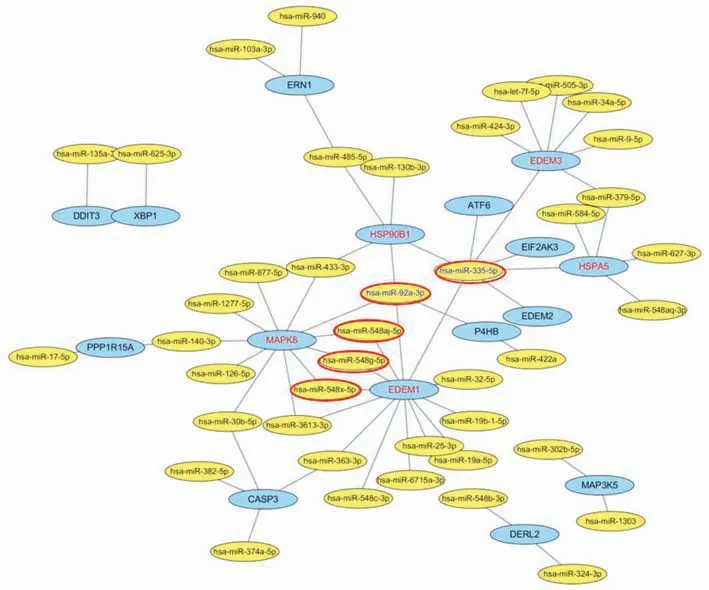
Fig.4 The difference of miRNA expression was obtained between human astrocytes and human neurons transfected with EIF2B5-RNAi after ERS stimulation by screening strategy
3 Discussion
The eIF2B subunit has guanine nucleotide exchange factor(GEF)activity.It is a key component for protein translation initiation.It is still unknown why the white matter selectively involved in VWM patients,while the neurons in the brain is spared.Motor regression is more serious and earlier than intellectual damage in VWM patients.In our previous study on mutant oligodendrocytes,the UPR pathway was overactivated after ERS,and autophagy pathway clearing unfolded proteins was damaged,leading to massive apoptotic cell death[10].Astrocytes constitute the main part of the white matter of the brain and play an essential role in nutritional support and signal transmission for oligodendrocytes.The eIF2Bε subunit encoded byEIF2B5,is the largest one and the main functional domain of GEF catalysis[14-15].Studies in zebrafish have shown that mutation inEIF2B5may aggravate the response to stress and impair motor behavior with the effects on myelination,loss of oligodendrocyte precursor cells,increased apoptosis in the CNS in a zebrafish withEIF2Bmutants[16].
The leukodystrophy may be due to defects in protein translation,as eIF2B is the important protein translation initiation factor.MiRNAs are non-coding RNAs with a length of 21-22 nt involved in posttranscriptional regulation of gene expression.In vivoevidence showed miRNA overexpressing in mice improved myelin thickness,and may enhance both oligodendrocyte differentiation and myelin synthesis[17-18].MiRNAs also participate in the pathogenesis of neurodegenerative disease by regulating astrocyte function through different mechanisms and may be potential treatment targets[19].MiRNAs also play an important role in regulating a large number of developmental processes and diseases through biological networks[20-21].
As miRNAs are potent regulators and have been shown to drive multiple pathways,the impact of miRNA-miRNA interactions could be profound.Strategies designed for new individual miRNAs have limited effects,and most of them act in clusters,not many miRNA studies have considered the wider impact of miRNA on overall miRNA expression[22].MiRNAs may exert function in groups.Sequence and functional analysisin vitrofor selecting the single piece of miRNA may not exhibit the whole pictures of its influence and individual miRNAs have limited effects.In our study,changes in the participating number of miRNAs were calculated.After detecting all known or unknown miRNAs with two-fold or more expression differences,it was found that more miRNAs were involved in the regulation of wild-type astrocytes after ERS stimulation,while fewer involved in the regulation of wild-type neurons after ERS stimulation.Astrocytes may be more dependent on miRNA regulation than neurons.However,whenEIF2B5gene was knocked down in astrocytes,the number of miRNAs involved in the regulation after ERS stimulation was significantly reduced,accompanied with increased cell apoptosis and decreased cell viability.A series of comparison strategies were designed for screening which miRNA participated after ERS.The changes of miRNAs in wild-type orEIF2B5knockdown neurons and astrocytes with or without ERS stimulation were compared.The database was then returned to look for changes originating from natural differences,loss of function inEIF2B5,or ERS stimulation resulting in differences.Cluster analysis was performed on the final miRNAs with differences to find those closely related to each known pathway.Five potential meaningful known miRNAs were obtained.These 5 miRNAs serve as junctions that connecting different pathways which involved in many biological processes as cell proliferation,differentiation,apoptosis,and maturation through gene repression.
4 Conclusion
Astrocytes and oligodendrocytes,both of which belong to the secretory pathway,may rely more on numerous miRNAs for the regulation of repairment to promote survival after ERS compared with neurons.EIF2B5-RNAi human astrocytes have a serious shortage of miRNAs participated after ERS due to the spontaneous apoptosis,which may lead to failure to survive in the last stage.Endogenous non-coding RNAs as miRNAs participate in cell proliferation,differentiation and maturation.Further studies on related miRNAs provide diagnostic,prognostic markers and chemotherapeutic targets in VWM.The calculation of participated miRNAs and selected miRNAs interference may serve as another observation marker besides cell apoptosis and vitality for the intervene of UPR pathway.

- 生物化學(xué)與生物物理進(jìn)展的其它文章
- 42℃熱療抑制U251細(xì)胞增殖并誘導(dǎo)凋亡*
- The Dynamic Changes and Clinical Significance of Serum Neuroglobin Levels in Patients With Acute Ischemic Stroke
- 組織蛋白酶B介導(dǎo)NLRP3小體在砷致小膠質(zhì)細(xì)胞炎癥激活中的作用*
- TMEM163變異致髓鞘形成低下患者隨訪兩例及iPSC構(gòu)建*
- 一個(gè)白質(zhì)消融性白質(zhì)腦病家系新基因突變及臨床表型研究
- 第五版《世界衛(wèi)生組織中樞神經(jīng)系統(tǒng)腫瘤分類指南》下成人膠質(zhì)瘤診斷及預(yù)后*