
Granulocyte-macrophage colony-stimulating factor protects mice against hepatocellular carcinoma by ameliorating intestinal dysbiosis and attenuating inflammation
2020-10-22 04:31:38YongNaWuLeiZhangTuoChenXunLiLiHongHeGuangXiuLiu
World Journal of Gastroenterology 2020年36期
Yong-Na Wu, Lei Zhang, Tuo Chen, Xun Li, Li-Hong He, Guang-Xiu Liu
Abstract
Key Words: Granulocyte-macrophage colony-stimulating factor; Microbiome; Inflammation; Hepatocellular carcinoma
INTRODUCTION
Hepatocellular carcinoma (HCC) is currently the third leading cause of cancer mortality[1]. There are approximately 466000 new cases of HCC in China each year, accounting for approximately 55% of the total cases worldwide. HCC has become an extremely serious public health problem in China due to its high incidence, poor prognosis, and high rate of postsurgical recurrence[2,3]. Unfortunately, lack of effective treatment for liver cancer further increases the overall burden on the patients. There is currently substantial research on the characterization of the immunology and biochemistry of the gut, which scarcely involve metabolic organs. Recent interest has been focused on important physiological functions of the gut, such as maintaining energy homeostasis[4].
In recent decades, researchers have focused on the important contributions of the gut microbiota to the key aspects of human health, notably metabolism and immunity[5,6]. The liver is the largest solid organ in our body; it processes 80% of the blood derived from the gastrointestinal tractviaoutflow from the portal vein[7]. A large number of resident immune cells within the liver modulate immune responses, promote liver repair and detoxification, and resist invasion of pathogens[8,9]. The gut microbiome is a critical factor that regulates liver immunity and maintains liver homeostasisviaa variety of mechanisms, including those involving endotoxin, Tolllike receptors (TLR), and bile acid metabolism[10-15]. Of particular interest, extensive studies have confirmed specific differences in the intestinal flora between patients with HCC and healthy control subjects. However, the specific mechanisms underlying these findings remain unclear[16,17].
Granulocyte–macrophage colony-stimulating factor (GM-CSF) is a host cytokine that stimulates bone marrow precursors to generate monocytes and macrophages. Furthermore, it enhances monocyte antibacterial and antitumor activities[18]. Microbiota can promote the release of GM-CSF; this process is of great importance in maintaining intestinal immune homeostasis. Interestingly, macrophage-mediated sensing of microbial signals controls GM-CSF levels[19]. Serum and tissue levels of GMCSF were decreased in patients with HCC. Conversely, we found that GM-CSF overexpression inhibited the proliferation, invasion, and migration of liver cancer cells and promoted their apoptosis (unpublished data). We hypothesize that GM-CSF plays a protective role against HCC by regulating intestinal microecology. In this study, we examined the effects of GM-CSF overexpression on the intestinal microecology and metabolism of HCC. Our findings demonstrated that GM-CSF overexpression protects mice against HCC by ameliorating intestinal dysbiosis, stabilizing the intestinal barrier, attenuating inflammation, and reducing serum endotoxin levels. These findings may have profound implications for the development of therapies for HCC.
MATERIALS AND METHODS
Ethics statement
This study was approved by the Ethics Committee of the First Hospital of Lanzhou University (No. LLYYLL-2017-18) and was performed in accordance with the Guidelines for Experimental Animals of the Ministry of Science and Technology (Beijing, China).
Animals and treatments
Seven-week-old male BALB/c nude mice were housed in a specific pathogen-free environment with temperature (23 ± 2°C) and humidity controls (approximately 50%) and a daily cycle of 12-h light/12-h dark together withad libitumaccess to food and drinking water. After 1 wk of acclimatization, the mice were randomly divided into three groups: Control (n= 10), HCC (n= 13), and HCC + GM-CSF (GM-CSF overexpression,n= 13) groups. In the HCC and HCC + GM-CSF groups, subcutaneous tumors were implanted in the early stage. Mouse orthotopic transplantation tumor models of HCC were established after subcutaneous tumors were successfully implanted. The specific operation method is as follows. The human GM-CSF sequence was transfected using lentivirus into the human HCC cell line HCCLM3. The cells growing in the log phase were injected into the armpit of nude mice under sterile conditions (1 × 107/mL). Then, the tumors were subcutaneously inoculated with cells for approximately 4 wks (approximately 1 cm in diameter, no rupture on the surface). The tumors were trimmed into a tumor mass of 1.5 mm × 2 mm × 1 mm, the abdominal wall was cut, and the right lobe of the liver was exposed and incised. The aforementioned spare tumor mass was implanted, followed by suturing and closing of the abdomen. After the effects of anesthesia wore off, the mice were returned to the cage for feeding. They were kept in this environment for 4 wk. The construction method of the HCC orthotopic tumor model was as described above.
Sample collection
Fecal samples were collected from all mice and stored in a -80°C freezer. Blood samples were collected and centrifuged at 3000 r/min for 15 min; all serum aliquots were stored at -80°C. Liver and colon samples were divided into halves; tissues from one part were fixed in 4% neutral buffered formaldehyde for paraffin embedding, whereas the other part was frozen in liquid nitrogen and stored at -80°C for subsequent RNA extraction.
Histological evaluation of liver and colon tissues
Paraffin-embedded liver and colon tissue sections were stained with hematoxylin and eosin (H&E) to observe the morphology of these tissues and to detect liver injury. Images were scanned using a NanoZoomer digital pathology system (Hamamatsu Photonics, K.K., Japan), which digitally scans the sections into a specific image format for further evaluation.
Serum parameter analysis
Alanine aminotransferase (ALT) and aspartate aminotransferase (AST) levels were measured using the ROCHE CABAS C311 analyzer (Roche, Germany). Lipopolysaccharide (LPS) concentrations as an indirect measure of endotoxin levels were measured with ELISA according to the manufacturer’s instructions (Guduo, Shanghai, China). Immune inflammatory factors were measured using commercial ELISA kits (Abbkine Scientific Co, United States).
Immunohistochemical staining
Liver and colon tissue sections were stained using an immunohistochemistry kit (DAKO, Denmark) as previously described[20]. The NanoZoomer digital pathology scanner was used to scan the stained sections into a format suitable for analysis. Each section was randomly selected at 200× and 400× magnifications for analysis. Image-Pro Plus software was used to measure the average optical density of tight junction protein-1 (ZO-1).
RNA extraction and real-time PCR analysis
Total RNA was extracted from liver and colon tissues using an RNA kit (Qiagen, Germantown, MD, United States) according to the manufacturer’s protocol. Real-time PCR was performed with the Applied Biosystems 7500 system using the one-step SYBR PrimeScript plus RT-PCR kit (Takara, Japan). The PCR primer sequences used in this study are listed in Supplementary Table S1. The expression data of the samples were normalized to those of the internal control glyceraldehyde 3-phosphate dehydrogenase (GAPDH) and were calculated according to the ΔΔCt method.
Analysis of microbial communities in fecal samples
The QIAamp Rapid DNA Kit (Qiagen) was used to extract DNA from mouse stool samples. The DNA concentration and integrity were measured using a NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, Hudson, NH, United States) and agarose gel electrophoresis, respectively. Suitable DNA samples were transported to Genesky Biotechnologies Inc., Shanghai, China, for absolute quantification and 16S rRNA gene amplicon sequencing. The spike-in sequences contained conserved regions identical to those of natural 16S rRNA genes and artificial variable regions and were distinct from those found in nucleotide sequences in public databases. These served as internal standards and facilitated absolute quantification across samples. Appropriate mixtures with known copy numbers of spike-in sequences were added to the sample DNAs. The V3-V4 regions of the 16S rRNA genes and spike-in sequences were PCRamplified and sequenced using Illumina HiSeq.
Metabolomic profiling of fecal samples
Metabolomics analysis was performed using an Agilent 7890 gas chromatography time-of-flight mass spectrometry (GC-TOF-MS). ChromaTOF software was used to analyze the mass spectrum data for peak extraction, baseline correction, deconvolution, peak integration, and peak alignment[21]. For specific identification of biomolecules, the LECO-Fiehn Rtx5 database was used to match mass spectra and the retention time index. Finally, the peaks with a detection rate below 50% or relative standard deviation (RSD) > 30% in the quality control (QC) samples were removed[22]. Metabolites were identified from the US National Institutes of Standards and Technology (NIST) and Fiehn databases. Orthogonal partial least squares-discriminant analysis (OPLS-DA) was performed to visualize metabolic differences between experimental groups. Differential detection of metabolites was based on the statistical significance (Pvalue) of the two-tailed Student’stteston the normalized peak areas, and metabolites withvariable importance in the projection values > 1 andPvalues < 0.05 were included.
Statistical analysis
The data are presented as the means ± SE. For most data, one-way ANOVA with Tukey’s post hoc test was used to determine statistically significant differences between the groups. The Wilcoxon rank-sum test was performed to evaluate alpha diversity and principal coordinates between the different cohorts in absolute quantificative 16S rRNA amplicon sequencing analysis. PERMANOVA (Adonis) was used to test microbial community clustering using both weighted and unweighted UniFrac and Bray-Curtis distance matrices. Correlations between the variables were determined using Spearman’s rank correlation;Pvalues < 0.05 were considered significant. The data were analyzed using SPSS version 19.0 for Windows (SPSS Inc., United States).
RESULTS
GM-CSF overexpression protected against hepatic injury, normalized liver function, stabilized the intestinal barrier, and reduced serum endotoxin levels secondary to HCC
To assess the effects of GM-CSF on hepatic injury, liver function, intestinal barrier function, and serum endotoxin levels, we established an orthotopic transplantation tumor model of HCC by transecting GM-CSF-encoded lentivirus into an HCC cell line. The morphology of the livers in the HCC + GM-CSF group improved compared with that in the mice with HCC alone; tumor tissue infiltration was also reduced in the HCC + GM-CSF group (Figure 1A). The normal structure of the liver of the mice in the HCC group was extensively destroyed by tumor tissue infiltration, and there was considerable accumulation of vesicular fat, which is related to mixed inflammatory infiltration. In contrast, GM-CSF showed no impact on colon morphology (Figure 1C).
The mice in the HCC group had significantly higher serum ALT and AST levels than the controls. However, GM-CSF overexpression in HCC cells suppressed serum ALT and AST levels (Figure 1E). Zhuet al[23]reported that damage to intestinal tight junctions was associated with loss of intestinal barrier function. To explore this possibility as well as to examine the distribution of ZO-1 in the liver and colon[24], we first confirmed the increase in the levels of ZO-1 protein in the HCC + GM-CSF groupviaimmunohistochemistry staining. These findings suggest that GM-CSF overexpression reversed intestinal barrier dysfunction observed in response to HCC (Figure 1B and D). In parallel, the expression of ZO-1 transcripts in the liver and colon was upregulated among mice in the HCC + GM-CSF group (Figure 1F).
Endotoxin activates specific Toll-like receptors[25]; serum LPS levels were significantly reduced in the mice in the HCC + GM-CSF group compared with those in the HCC alone group (Figure 1G). Taken together, these results suggest that GM-CSF overexpression alleviated hepatic injury, normalized liver function tests, stabilized the intestinal barrier, and reduced serum endotoxin levels observed in response to HCC.
GM-CSF overexpression resulted in normalized cytokine profiles and Toll-like receptor expression
Current research indicates that cytokines play an important role in the progression of HCC[26]. GM-CSF promoted significant downregulation of proinflammatory cytokines [interleukin (IL)-1β and IL-2] and increased the levels of anti-inflammatory cytokines IL-4 and IL-10, with no effects on tumor necrosis factor levels. To explore and extend these findings with respect to our mouse model, we measured the expression of cytokine transcripts in the liver tissue. GM-CSF overexpression resulted in a significant decrease in the expression of IL-1β and TGFβ; however, no changes were observed with respect to IL-4 expression (Figure 2B). Furthermore, we found that GMCSF overexpression resulted in increased IL-4 expression in the colon, whereas there were no changes in IL-1β expression (Figure 2C). Toll-like receptor (TLR)-4 and its coreceptor CD14 are central to LPS recognition[27]. Endotoxin-associated increases in both TLR4 and CD14 expression in the liver and colon were reversed by GM-CSF. These results suggest the direct effects of GM-CSF on modulating the expression of both TLR4 and CD14.
GM-CSF ameliorated intestinal dysbiosis induced by HCC
To obtain a greater understanding of the protective effects of GM-CSF, we examined the intestinal microbiome of the three study groups using the absolute quantificative 16S rRNA amplicon sequencing technology with an artificially synthesized DNA internal standard; use of this standard permits us to simultaneously measure the absolute and relative abundance of specific fecal bacterial communities. While previous studies have reported the relative abundance of microorganisms in specific fecal samples, increasing concern has been focused on determining the absolute abundance of targeted micro-organisms in different samples.
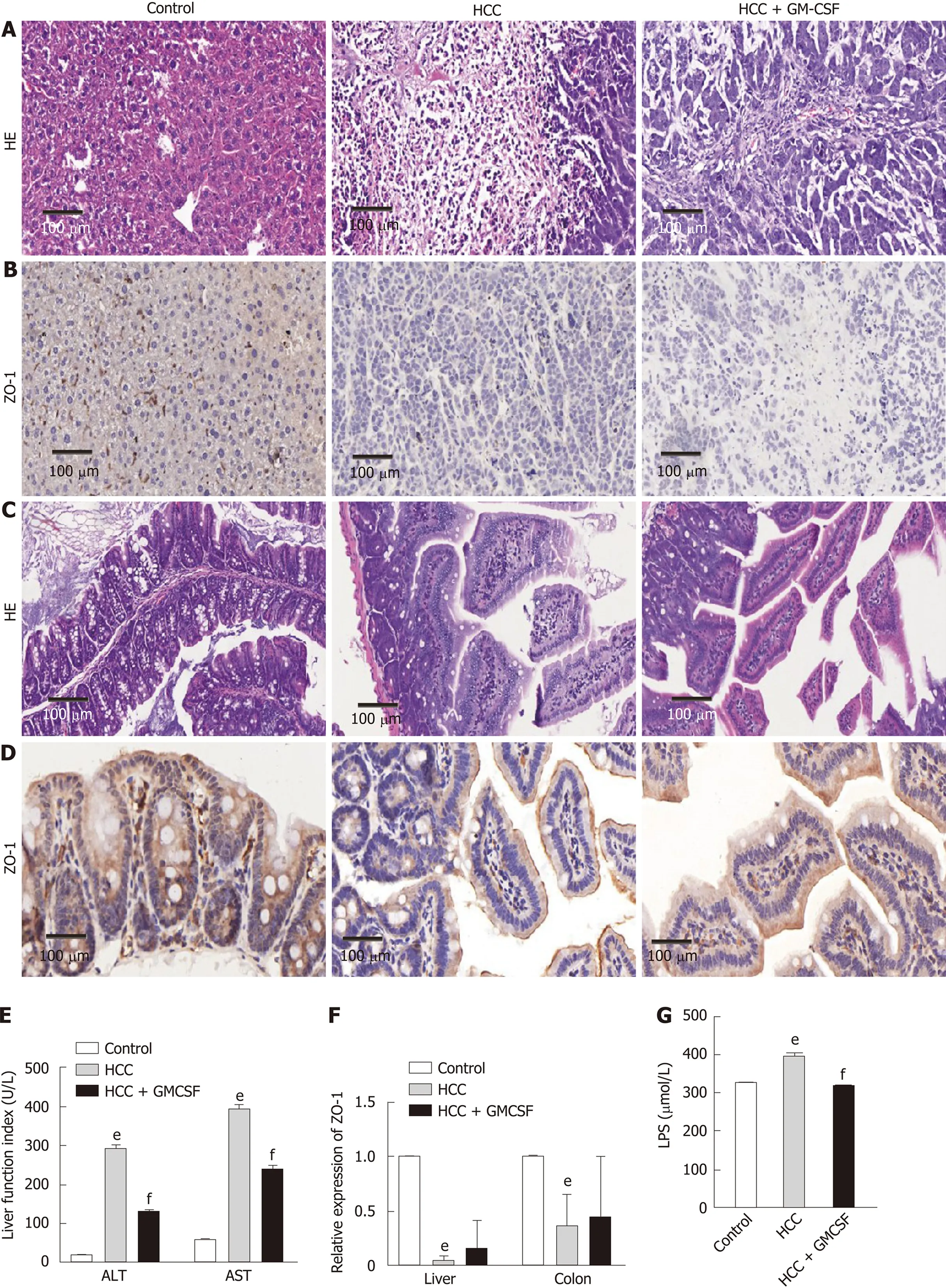
Figure 1 Granulocyte-macrophage colony-stimulating factor alleviated hepatic injury, normalized liver function, improved intestinal barrier function, and reduced serum endotoxin levels detected in response to hepatocellular carcinoma. A: Liver histology was assessed by hematoxylin and eosin (H&E) staining, scale bar: 200 μm (200 ×); B: Liver histology was assessed via ZO-1 immunohistochemical staining (scale bar, 200 μm); C: Colon histology was assessed via H&E staining, scale bar: 200 μm (400 ×); D: Colon histology was assessed via ZO-1 immunohistochemical staining (scale bar, 200 μm); E: Serum alanine aminotransferase and aspartate aminotransferase (U/L) levels; F: Expression of the intestinal barrier function marker ZO-1 in the liver and colon; G: Serum lipopolysaccharide levels (μmol/L). Data are presented as the means ± SE, n = 11–13 mice per group detected via one-way ANOVA with post hoc Tukey’s test. aP < 0.05, bP < 0.01, and eP < 0.001, HCC vs control; cP < 0.05, dP < 0.01, and fP < 0.001, HCC + GM-CSF vs HCC. HCC: Hepatocellular carcinoma; GM-CSF: Granulocyte-macrophage colony-stimulating factor; ALT: Alanine aminotransferase; AST: Aspartate aminotransferase; LPS: Lipopolysaccharide; H&E: Hematoxylin and eosin.
Alpha diversities were calculated using diversity indexes. Our results showed that the Chao index and number of operational taxonomic units were significantly increased in the fecal samples of HCC + GM-CSF mice; however, there were no significant differences in the Shannon index (Supplementary Figure 1A). Βeta diversity values were calculated by weighted UniFrac principal coordinates analysis to assess the phylogenetic relationship between microbial communities. The results from the control, HCC, and HCC + GM-CSF groups can be clearly separated into different clusters (Supplementary Figure 1C). Similarly, the structure of the intestinal flora of the HCC and HCC + GM-CSF groups was clearly divided into distinct clusters.
The relative abundance and absolute copy number of each bacterial group at the phylum level are shown in Figure 3A. The microbial composition of the HCC + GMCSF group was significantly different from that of the HCC alone group. Of note, the absolute abundance of the micro-organisms in the HCC group was reduced by 12% compared with the control group. Furthermore, the absolute abundance of intestinal microorganisms among the fecal samples from the HCC + GM-CSF group was 1.35 times higher than among those in the HCC group; these findings suggest that GM-CSF limits HCC-induced intestinal dysbiosis. The intestinal microbiota identified in all three groups primarily included bacteria within the phyla Firmicutes, Bacteroidetes, Deferribacteres, Tenericutes, Proteobacteria, Actinobacteria, and Cyanobacteria (Figure 3A). Among them, the abundance of Tenericutes, which is related to hepatopancreatic necrosis, was 2.45 times higher in the samples from the HCC groups than those from the control. GM-CSF overexpression resulted in a 63% decrease in the absolute abundance of Tenericutes compared with that observed in samples from mice with HCC. At the genus level (Figure 3B), the intestinal flora mainly includes
Parabacteroides,Prevotella,Anaerobacterium,Eubacterium,Clostridium_lV,Anaerotruncus,Mucispirillum,Rosecusburia, andButyricicoccus. We analyzed the absolute abundance at the genus level as shown in Figure 4.
To evaluate the differences in the intestinal microbiome at the bacterial genus level, we performed statistical evaluation focusing on the most prominent differences among the three groups. GM-CSF overexpression contributed to the high abundance of protective genera (e.g.,Roseburia,Blautia, andButyricimonass), along with a significant reduction inPrevotella,Parabacteroides,Anaerotruncu,Streptococcus,Clostridium, and
Mucispirillum. Particularly notable was the genusPrevotella, which was reduced 10 000-fold in response to GM-CSF overexpression. To identify additional differences in the intestinal flora among the different groups, we performed the linear discriminant analysis effect size (LEfse) analysis based on the ribosomal database project classification data (Supplementary Figures 2C and D) and identified the most important differences between the HCC and HCC + GM-CSF groups, including those identified among thePrevotella,Clostridium, andAnaerotruncusgenera.
GM-CSF normalization of the fecal metabolomic profile
We further elucidated the intestinal dysfunction secondary to HCC through a nontargeted metabolomics study. A total of 305 metabolites included in the following Kyoto Encyclopedia of Genes and Genomes (KEGG) metabolic pathways were identified: Biotin (vitamin) anabolic, sphingolipid, steroid anabolic, pyrimidine, citric acid cycle (TCA cycle), purine, and primary bile acid metabolism (Figure 5C). A corresponding OPLS-DA analysis was performed. As shown in Figure 5A, the score graph reveals that the metabolomic profiles from each group are clearly clustered. We further established the OPLS-DA model corresponding to the HCC and HCC + GMCSF groups; the results demonstrated that the HCC and HCC + GM-CSF groups could be clearly divided (Figure 5B). A total of 69 metabolites were screened; we identified significantly higher concentrations of biotin and oleic acid in the HCC + GM-CSF group. These two metabolites were involved in biotin and lipid metabolism, respectively. Likewise, succinic acid, fumaric acid, adenosine, and maleic acid were detected at significantly reduced concentrations in samples from the HCC + GM-CSF group (Figure 5D).
GM-CSF remodeling of intestinal microecology and function
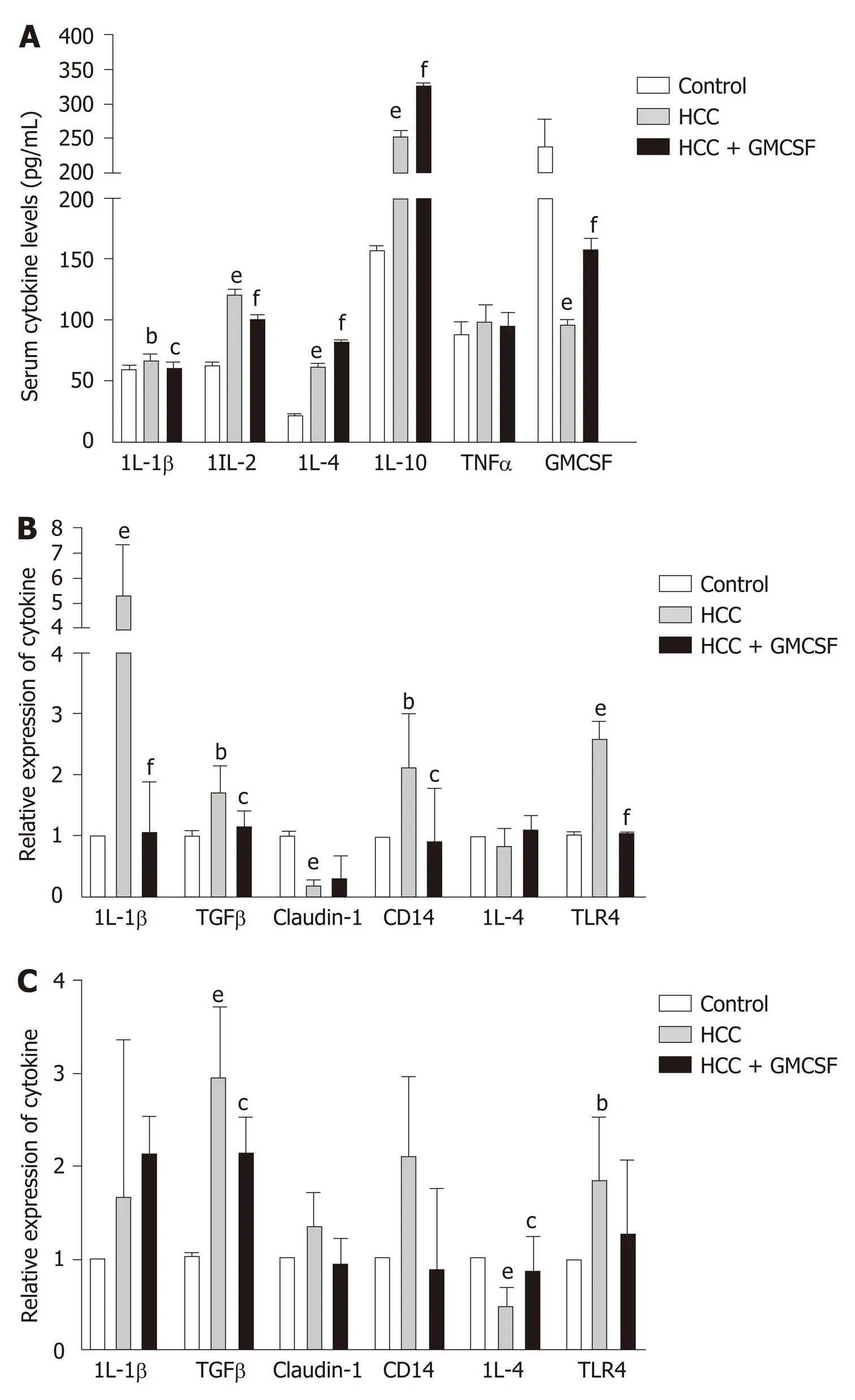
Figure 2 Granulocyte-macrophage colony-stimulating factor normalized cytokine profiles and regulated Toll-like receptor expression. A: Cytokine levels in serum; B and C: Cytokine gene expression in the liver and colon. Data are presented as the means ± SE and analyzed via one-way ANOVA with post hoc Tukey’s test. n = 11-13 mice per group. aP < 0.05, bP < 0.01, and eP < 0.001 hepatocellular carcinoma (HCC) vs control; cP < 0.05, dP < 0.01, and fP < 0.001 HCC + GM-CSF vs HCC. IL-1β: Interleukin-1β; TGFβ: Transforming growth factor β; HCC: Hepatocellular carcinoma; GM-CSF: Granulocyte-macrophage colonystimulating factor.
To determine the beneficial impact of GM-CSF on intestinal microecology, we conducted a correlation analysis of the interaction matrix. As part of the analysis, we defined the following: Representative microbial genera with significant alterations in abundance in the HCC and HCC + GM-CSF groups, metabolites detected at significantly different concentrations in the aforementioned two groups, and other representative parameters related to liver and intestine function (Figure 6). GM-CSF overexpression can promote remodeling of the intestinal microecology and function resulting from the development of HCC. A significant reduction in the abundance ofPrevotellain the HCC + GM-CSF group positively correlated with serum ALT and LPS levels and inversely correlated with serum IL-10 and GM-CSF levels. Additionally, the abundance of bacteria of thePrevotellagenus positively correlated with succinic acid levels, with the latter being significantly higher in fecal samples from the HCC group. Similarly, the abundance of the bacterial genusAnaerotruncus, which may be involved in the inflammatory response, was strongly positively correlated with serum ALT levels and inversely correlated with ZO-1 levels and IL-4 expression. Biotin levels, which were high in samples in response to GM-CSF overexpression, inversely correlated with IL-2 expression. Likewise, succinic acid, which was detected at relatively low levels in the fecal samples of mice in the HCC + GM-CSF group, positively correlated with ALT, AST, and LPS levels and inversely correlated with IL-4 and IL-10 expression. Taken together, these data provide evidence that GM-CSF plays a dominant role in remodeling the aberrant intestinal microecology and function that develop in response to HCC.
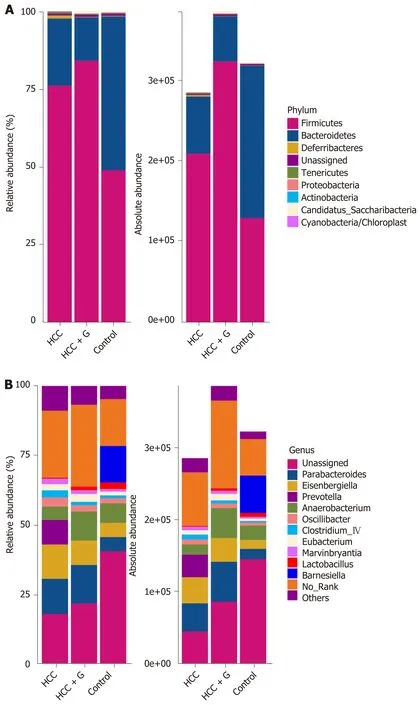
Figure 3 Relative and absolute abundances of the major bacteria at phylum (A) and genus (B) levels.
DISCUSSION
This study demonstrated that GM-CSF overexpression can repair intestinal dysbiosis, liver damage, inflammation, and damage to liver function induced by HCC. Specifically, several complementary GM-CSF-mediated mechanisms have been revealed, including a role in (1) protecting the intestinal barrier; (2) regulating biotin and lipid metabolism; (3) attenuating inflammation; and (4) reducing endotoxin levels.
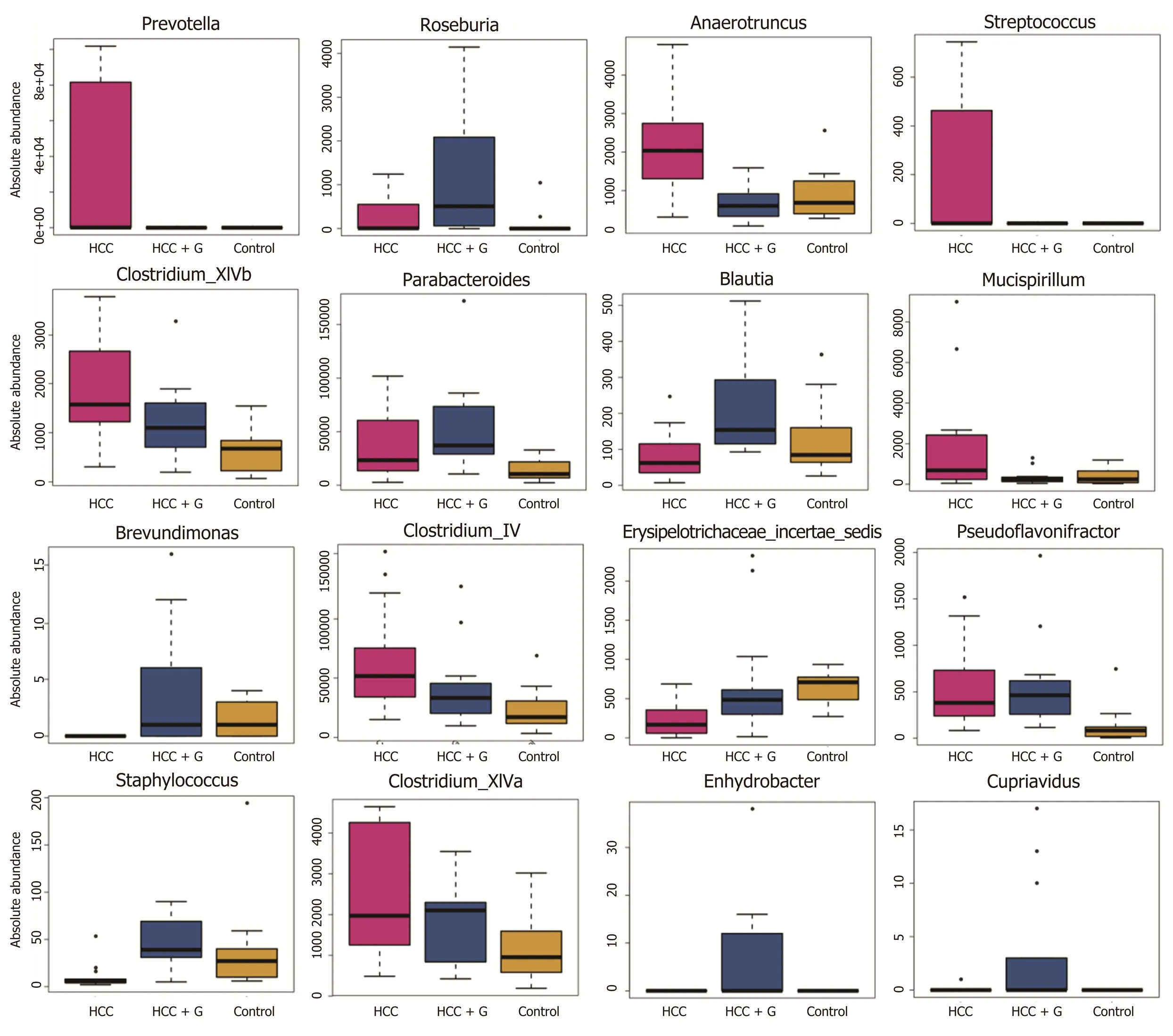
Figure 4 Profiles of the gut microbes among the hepatocellular carcinoma + granulocyte-macrophage colony-stimulating factor, hepatocellular carcinoma and control groups. The absolute abundance of each genus is indicated on the y-axis. The ordinate represents the abundance value among the three groups. The Kruskal-Wallis rank-sum test was used to analyze the different species. The figure shows all the significantly different genera in the diversity indexes among the groups, and a P value < 0.05 was considered significantly different. HCC: Hepatocellular carcinoma; GM-CSF: Granulocytemacrophage colony-stimulating factor.
Enterohepatic recycling depends heavily on the nature and function of the intestinal micro-organisms; the intestinal flora also plays a critical role in protecting against infections[28]. TLR signaling triggers the initial inflammatory response by releasing cytokines, which may promote intestinal dysbiosis along with secondary TLR activation and signaling[29]. In this study, we found that GM-CSF overexpression resulted in significant downregulation of TLR4 expression in both liver and colon tissues. These findings suggest that GM-CSF provides protection from HCC-mediated destruction of the intestinal barrier and limits the inflammatory response secondary to LPS-mediated TLR4 signaling. The gut microbiota and metabolic products are translocated to the liverviathe portal circulation, consequently activating TLR4 in the immune cells localized in the hepatic tissue. Emerging studies have shown that intestinal mucosal damage and epithelial barrier disruption result in bacterial translocation and overgrowth; as such, endotoxins can accumulate in the liver. As the liver loses its toxin clearing ability, endotoxins will find their way into the systemic circulation[30]. This scenario has been reported in patients with alcoholic fatty liver in which significant elevations in systemic LPS have been detected[13]. In this study, we found that LPS levels were significantly elevated in the serum of mice with HCC; these levels were markedly reduced in mice with HCC and GM-CSF overexpression. Recent studies have shown that elevated LPS levels in patients with liver injury are mainly due to impaired intestinal barrier function and intestinal dysbiosis[31]. LPS is a pathogen-associated molecular pattern molecule that can trigger severe inflammatory responses both directly and indirectly[14]. Taken together, our data suggest that GMCSF may limit LPS-mediated activation of the TLR4 signaling pathway, thereby protecting against the inflammatory mic-roenvironment generated in response to HCC.
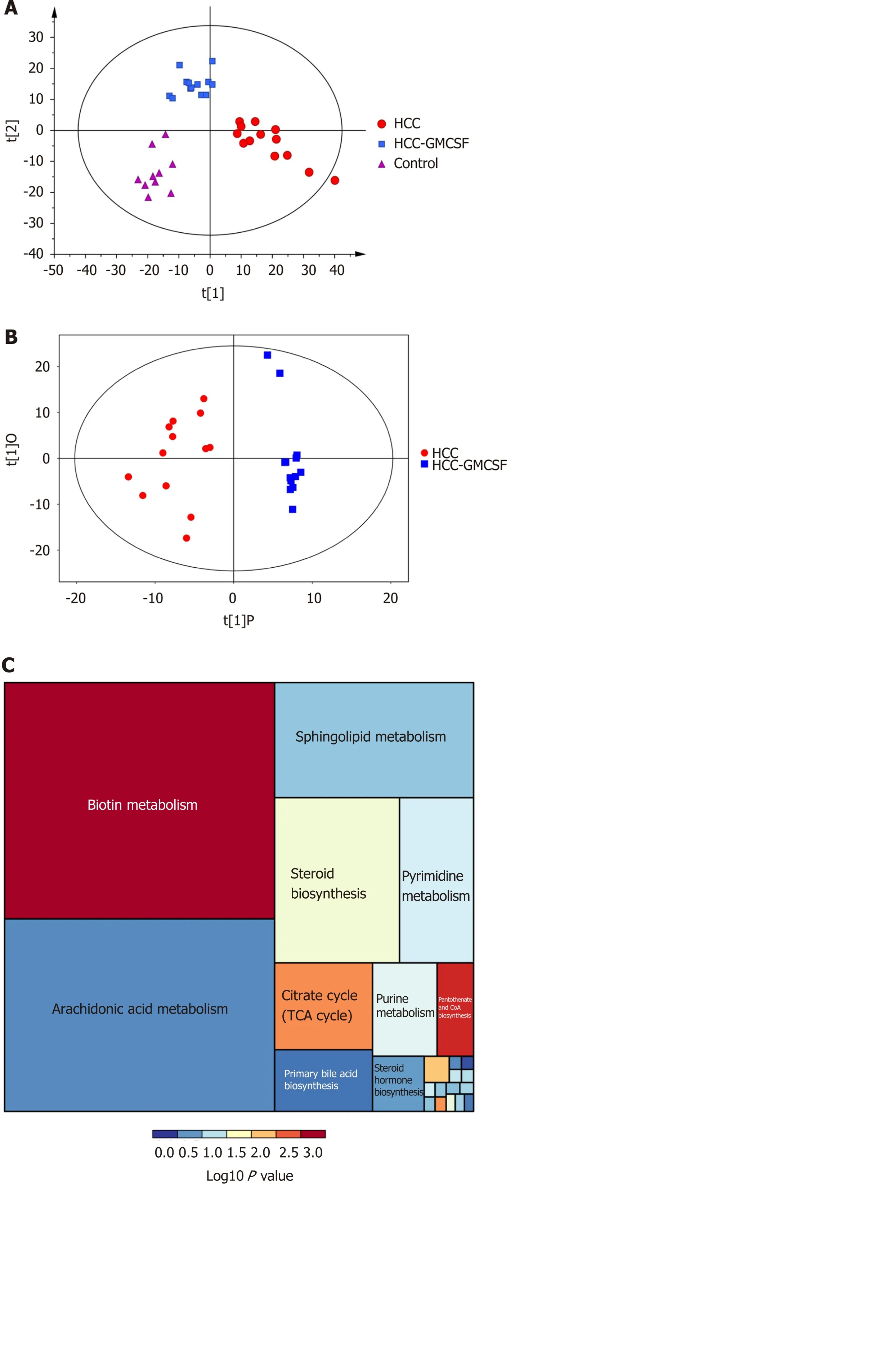
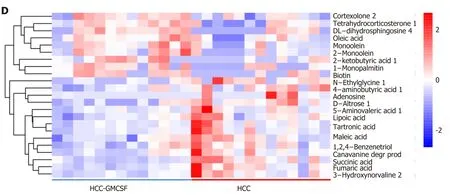
Figure 5 Granulocyte-macrophage colony-stimulating factor normalized the fecal metabolomic profile detected in response to hepatocellular carcinoma. A: OPLS-DA score scatter plot comparing the control (purple), hepatocellular carcinoma (HCC) (red), and HCC + granulocytemacrophage colony-stimulating factor (GM-CSF) (blue) groups; B: OPLS-DA score scatter plot comparing the HCC (red) and HCC + GM-CSF (blue) groups; C: Differential involvement of the metabolic pathways between the three groups; D: Heat map detailing the differential detection of metabolites comparing the HCC and HCC + GM-CSF groups. HCC: Hepatocellular carcinoma; GM-CSF: Granulocyte-macrophage colony-stimulating factor.
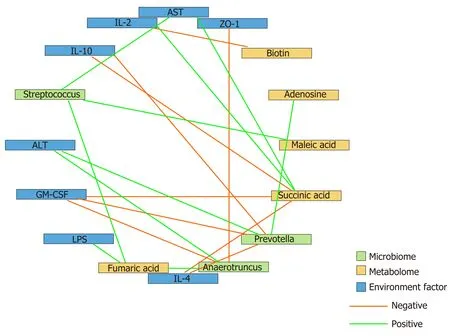
Figure 6 Correlation analysis of the representative microbial genera, characterized metabolites, and environmental factors (parameters indicating liver and gut impairment) among the control, hepatocellular carcinoma, and hepatocellular carcinoma + granulocytemacrophage colony-stimulating factor groups. Spearman’s rank correlation was used, and significant associations with P < 0.05 and r > 0.5 were found. Blue nodes, representative injury parameters; green nodes, differentially distributed genera; yellow nodes, metabolites with a high discriminative power between the groups. Green lines between nodes indicate positive linkages, and orange lines represent negative relationships. The thickness of the connection represents the correlation coefficient, with thicker lines indicating higher r values. ALT: Alanine aminotransferase; AST: Aspartate aminotransferase; LPS: Lipopolysaccharide; IL-2: Interleukin-2; GM-CSF: Granulocyte-macrophage colony-stimulating factor.
Previous studies have revealed thatBacillus subtilispromotes the expression of the tight junction protein ZO-1, alters intestinal immune activity, and affects the integrity of the intestinal barrier[32,33]. In this study, we detected ZO-1 expressionviaimmunohistochemistry and gene transcription analysis, and the results revealed that mice with HCC experienced intestinal barrier dysfunction, as indicated by a significant decrease in ZO-1 expression. Conversely, GM-CSF resulted in an increase in ZO-1 expression. Overall, our results indicate that GM-CSF overexpression protects against HCC-mediated disruption of intestinal barrier function.
Tumor growth typically takes place in a complex environment with cytokines, chemokines, and extracellular matrix components, which are collectively considered to be the “tumor microenvironment”[34]. HCC is a typical inflammation-related tumor; its initiation and development are closely associated with a specific tumor microenvironment[35]. Pollardet al[36]reported that macrophages in the tumor microenvironment are polarized into M1 and M2 phenotypes in response to GM-CSF. Macrophages can be recruited to tumor tissues where they can modulate and regulate the local immune response[37]. In this study, we found that liver and systemic inflammatory responses were ameliorated in response to GM-CSF overexpression; these observations may be directly related to the impact of GM-CSF on macrophage activation. In addition, we found that serum levels of the proinflammatory cytokines IL-1β and IL-2 and the liver enzymes ALT and AST were significantly downregulated in response to GM-CSF overexpression, whereas the expression of the antiinflammatory cytokines IL-4 and IL-10 was increased. These findings suggest that liver function was protected by GM-CSF overexpression and that liver cell damage and the associated inflammation were reduced.
A previous study has shown that translocation of intestinal flora results in chronic inflammation in the liver, thereby exacerbating HCC[38]. To generate an accurate evaluation of the structural characteristics of the intestinal flora, we determined the absolute copy number of the 16S rRNA genes in fecal samples from control mice and mice with HCC with or without GM-CSF overexpression. We identified an increase in the overall abundance of Tenericutes associated with hepatopancreatic necrosis disease in mice with HCC. In contrast, GM-CSF overexpression in association with HCC led to a 63% reduction in the overall abundance of bacteria of this phylum. It is also worth noting that GM-CSF overexpression in association with HCC resulted in a reduction in the overall abundance ofParabacteroide, Prevotella,Streptococcus, andClostridiumspp. and increased the overall abundance of anti-inflammatory bacterial species, includingBlautia,Butyricimonass, andRoseburia[39-41].
Recent research findings have suggested that patients with type 2 diabetes mellitus, inflammatory bowel diseases, colorectal adenocarcinoma, and liver cancer present with moderate dysbiosis, including a net decrease in the anti-inflammatory bacterial genera and an increase in various potential pathogens[42-45]. These results parallel our findings on the beneficial effects of GM-CSF overexpression in association with HCC.
In our previous research, we found an increase in the abundance ofPrevotellain the feces of patients with HCC compared with healthy volunteers[17].Prevotellais a conditional pathogen that was originally identified in the oral cavity and vagina;Prevotellaspp. is associated with periodontal disease. The identification of a large number of these bacteria in the feces of patients with HCC suggests translocation of oral bacteria to the gastrointestinal tract. The bacterial genomes ofPrevotellaspp. encode superoxide reductase, which facilitates resistance to host-derived reactive oxygen species and promotes intestinal inflammation by generating thioredoxin[46]. In this study, we found thatPrevotellapositively correlated with ALT, IL-2, and LPS levels and inversely correlated with IL-10 and GM-CSF; this finding is consistent with the conclusions from the aforementioned previous studies. Absolute numbers of bacteria from the phylumAnaerotruncuswere also present in significantly reduced numbers in the fecal samples of mice overexpressing GM-CSF; these bacteria may promote both intestinal and hepatic inflammatory responses.Anaerotruncuslevels strongly positively correlated with serum ALT levels and inversely correlated with the expression of the intestinal barrier function protein ZO-1 and the anti-inflammatory factor IL-4. Overall, these results imply that bacterial populations responding significantly to GM-CSF overexpression may be those regulating inflammation, immunity, and barrier functions.
Microbes are connected to each otherviacomplex metabolic chains. Studying microbial metabolism in the intestines may help in determining the interactions among the members of the microbial community. Therefore, to achieve a better understanding of the protective effects of GM-CSF against HCC, we integrated metabolomics into this study. We found that GM-CSF overexpression was associated with a significant increase in biotin and oleic acid levels and a concomitant decrease in succinic acid, adenosine, fumaric acid, lipoic acid, and maleic acid levels. As a group, these metabolites are closely related to biotin and lipid metabolism pathways. Biotin is a vitamin synthesized by intestinal microorganisms[47]that enhanced expression of glucokinase in rat liver[48]. When biotin levels are low, protein synthesis in the liver and intestinal mucosa is inhibited; protein synthesis is restored after biotin supplementation[49]. Likewise, succinic acid levels were diminished in the fecal samples of GM-CSF-overexpressing mice. Earlier studies have reported that succinic acid is a toxic factor directly associated with inflammation and disease[50]. However, no changes in bile acids were detected in our study. This may be because our research used a GCMS detection platform to collect data and the Fiehn and National Institute of Standards and Technology (NIST) databases to determine the nature of the substances. Because the database of the GC platform itself contains relatively few bile acids, there are no qualitative bile acids available. In the future, the LC platform will be used to detect related bile acid substances, and relevant research will be conducted. We found that succinic acid levels positively correlated with serum ALT, AST, and LPS levels and inversely correlated with IL-4 and IL-10 levels; these findings are consistent with previous results. Interestingly,Prevotellaare succinic acid-producing bacteria, explaining the positive correlation between succinic acid andPrevotella. GM-CSF overexpression resulted in the reduced abundance ofPrevotella[51], possibly constituting an effective feedback loop in the intestinal microecosystem, serving to magnify the protective effects of GM-CSF overexpression.
CONCLUSION
In summary, we have demonstrated that GM-CSF overexpression can ameliorate intestinal dysbiosis, attenuate inflammation, and reduce endotoxin levels associated with HCC pathogenesis. In light of our findings, a more comprehensive understanding of the impact of GM-CSF in this setting might lead to new insights into the treatment for HCC.
ARTICLE HIGHLIGHTS
Research background
Granulocyte-macrophage colony-stimulating factor (GM-CSF) plays a contributing role in the pathogenesis of hepatocellular carcinoma (HCC) progression, and GM-CSF is a critical factor in promoting intestinal immunity.
Research motivation
GM-CSF may protect against the development of HCC by regulating immunity as well as the intestinal microecology.
Research objectives
To investigate the impact of GM-CSF on the gut microbiome and metabolic characteristics of HCC.
Research methods
We utilized established mouse models of HCC and overexpressed GM-CSF in HCC. Liver injury, intestinal barrier function, immune inflammation and the fecal microbiome and metabolome were studied.
Research results
Overexpression of GM-CSF had a significant impact on the gut microbiome of mice with HCC and resulted in a high abundance of organisms from the generaRoseburia,BlautiaandButyricimonass, along with a significant reduction inPrevotella,Parabacteroides,Anaerotruncus,Streptococcus,Clostridium, andMucispirillum. GM-CSF overexpression resulted in a substantial increase in fecal biotin and oleic acid, along with a prominent decrease in the fecal levels of succinic acid, adenosine, fumaric acid, lipoic acid, and maleic acid. The intestinal microbiota and fecal metabolites induced by GM-CSF are primarily involved in the pathways related to the reduction in the inflammatory response, biotin metabolism and intestinal barrier dysfunction.
Research conclusions
GM-CSF protects mice against HCC by ameliorating intestinal dysbiosis and attenuating inflammation.
Research perspectives
Our findings indicate that GM-CSF can protect against the development of HCC by regulating immunity as well as by modulating the abundance of specific intestinal micro-organisms and their metabolites. This study provides new insights into the treatment of HCC.
 World Journal of Gastroenterology2020年36期
World Journal of Gastroenterology2020年36期
- World Journal of Gastroenterology的其它文章
- Pediatric non-alcoholic fatty liver disease and kidney function: Effect of HSD17B13 variant
- Major gastrointestinal bleeding and antithrombotics: Characteristics and management
- Artificial intelligence in gastric cancer: Application and future perspectives
- Treatment of eosinophlic esophagitis with swallowed topical corticosteroids
- SMARCB1/INI1-deficient pancreatic undifferentiated rhabdoid carcinoma mimicking solid pseudopapillary neoplasm: A case report and review of the literature
- Solitary peritoneal metastasis of gastrointestinal stromal tumor: A case report