
Calcareous nannoplankton dating of the Late Quaternary deposits in Greece and the eastern Mediterranean:Case studies from terrestrial and marine sites
2015-08-20 03:02:44MariaTriantaphyllou
Journal of Palaeogeography 2015年4期
Maria V.Triantaphyllou
Faculty of Geology and Geoenvironment,University of Athens,Panepistimioupolis Zografou,157-84 Athens,Greece
Keywords:Emiliania huxleyi Biostratigraphy Palaeoclimatic conditions Greece Eastern Mediterranean
ABSTRACT The distribution and abundance of Emiliania huxleyi(E.huxleyi)assemblages in the marine sediments of the Aravonitsa Plateau,Greece,and from the eastern Mediterranean are used(1)to evaluate the calcareous nannoplankton NN21a and NN21b biozones and the NN21a/NN21b boundary,and(2)to analyze the palaeoenvironmental and palaeoclimatic conditions prevailing in this interval.The sediment succession displays varied E.huxleyi assemblages and these are interpreted as reflecting climatic variability during marine isotope stages MIS 1-8.
1.Introduction
Emiliania huxleyi(E.huxleyi)is by far the most abundant and widespread coccolithophore in the world's oceans(Winter et al.,1994;Young,1994).It was first identified using light microscopy(Lohmann,1902)whereas the complicated structure of the coccolith was initially described by Black and Barnes(1961),Braarud et al.(1952),and Deflandre and Fert(1954),using Electron Microscopy.Three types(A,B and C)of E.huxleyi are distinguished(Young and Westbroek,1991),based on heterococcolith morphology,and it was demonstrated that the size of the different morphotypes is in fluenced by ecophenotypic and genotypic factors.The phylogenetic origins of E.huxleyi are documented in the fossil recordby Gallagher(1989),Perch-Nielsen(1985),Romein(1979)and Young et al.(1992).The monospecific genus Emiliania is considered to have evolved from the genus Prinsius through Toweius,Reticulofenestra and Gephyrocapsa(Perch-Nielsen,1985;Samtleben,1980;Young et al.,1992).
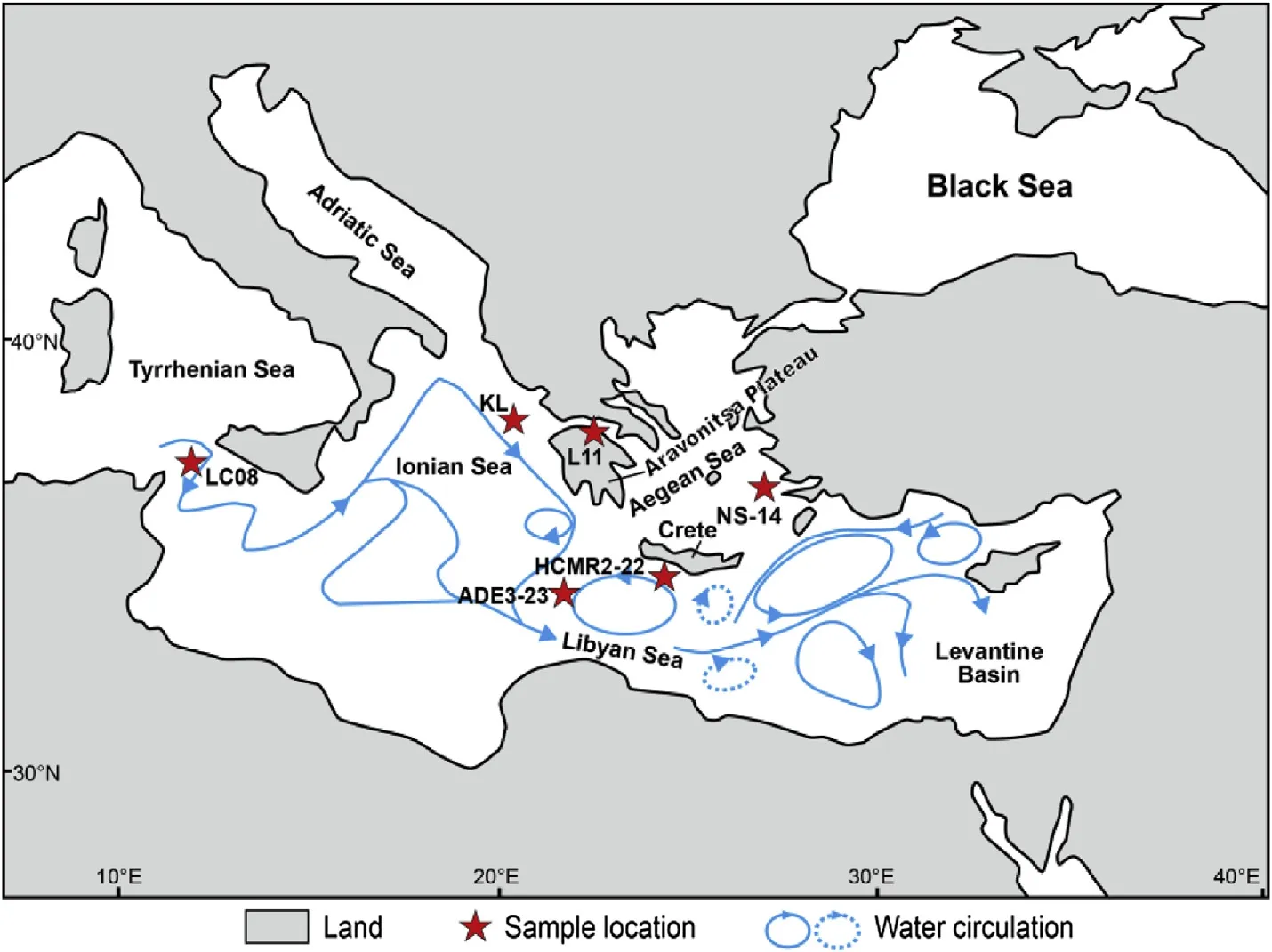
Fig.1-Location of the reviewed on-land and marine records in the eastern Mediterranean.The general circulation pattern is modified from Malanotte-Rizzoli et al.(1997).
Amongst the extant coccolithophores,E.huxleyi has the widest distribution,dominating the living assemblages.It occasionally forms massive blooms,when water conditions are favorable,e.g.high light,limited silicate and high carbonate saturation(Tyrrell and Merico,2004).Under certain environmental conditions it sheds coccoliths(Paasche,2001).Variation in the coccolith size and morphology of E.huxleyi is frequent(Young and Westbroek,1991),being usually associated with temperature,salinity and available nutrients(e.g.,Bollmann and Herrle,2007;Paasche,1998,2001;Watabe and Wilbur,1966).The increase in atmospheric CO2partial pressure and the consequent changes in the seawater carbonate chemistry can cause a decrease in its cellular particulate inorganic carbon(PIC)/particulate organic carbon(POC)ratio as well as malformations of the coccoliths(Riebesell et al.,2000)-these might have opposing effects on the marine carbon cycle (Rostand Riebesell,2004).Recently,the morphological variability of E.huxleyi var.huxleyi(=E.huxleyi type A)has been demonstrated in the modern Aegean Sea,in respectto theenvironmental parametersof the watercolumn,providing strong evidence for seasonal variation in E.huxleyi coccoliths(Triantaphyllou et al.,2010b).A consistent pattern of increase in the size and calcification of coccoliths and coccospheres has been observed,including the thickness of the inner tube elements in the winter and spring and low seasurface temperature and moderate productivity when compared with summer time,high temperature and low productivity(Triantaphyllou et al.,2010b).
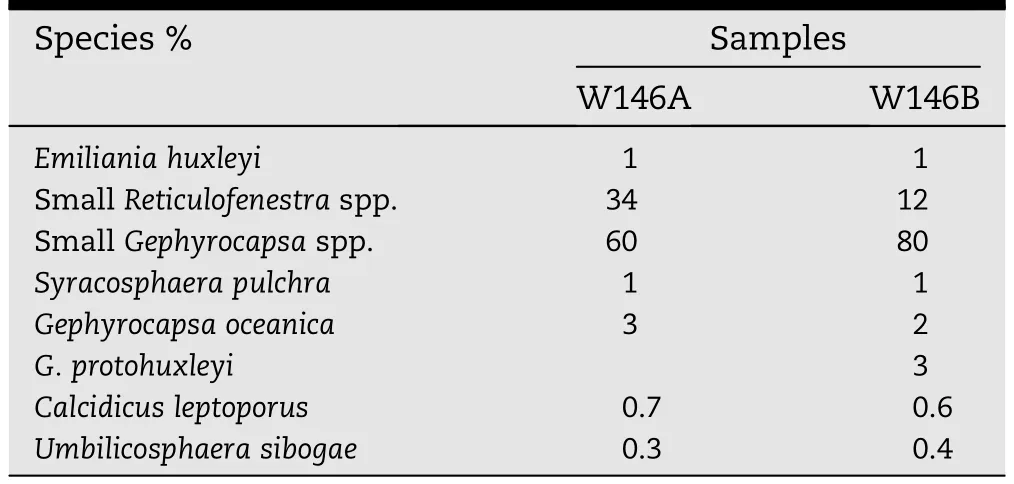
Table 1-Aravonitsa samples:Quantitative SEM analysis of 300 coccoliths.
The first appearance datum(FAD)of E.huxleyi in the geological records is used as a biostratigraphic marker and marks the base of the NN21 calcareous nannofossil biozone(NN21a;Martini,1971).It has been dated at~270 ka using correlation with planktonicforaminiferalδ18O records(Gartner and Emiliani,1976),at~268 ka,late in the cold marine isotopic stage 8(MIS 8;Thierstein et al.,1977),at~285 ka(Pacific ocean;Ahagon et al.,1993),and at~230 ka(Hills and Thierstein,1989).According to Kroon et al.(1998),this event is placed at 260 ka in the eastern Mediterranean,whereas the astronomically tuned biozone timescale of Lourens et al.(2005)dates the lowest occurrence of E.huxleyi and subsequently the base of the calcareous nannofossil biozone MNN21a(Raf fi,2002;Rio et al.,1990),(=NN21a;Martini,1971)at 270 ka(265 ka according to Raf fiet al.,2006).
The blooming life-style of E.huxleyi started in warm timespans between~80 ka at the end of MIS 5 and 50 ka(MIS 3)(Kroon et al.,1998),and is associated with the E.huxleyi Acme Zone(NN21b;Martini,1971)in the fossil record.In shallow marginaland inlandseas theE.huxleyiAcmeZone beganlater;in these areas Gephyrocapsa spp.are generally more dominant(e.g.,Okadaand Honjo,1975).The base of NN21b is followed by the top of the Gephyrocapsa muellerae Acme Zone at 45.7 ka(Flores et al.,1997;Incarbona et al.,2009),characterized by a dramatic and continuous reduction in G.muellerae,which suggests that E.huxleyi may have taken over an ecological niche formerly dominated by Gephyrocapsa spp.

Fig.2-A-E-Gephyrocapsa protohuxleyi specimens possessing a bridge or the remains of the bases of a bridge;F-G-Small Reticulofenestra specimens that have undergone etching resulting in separation of the ends of the elements(arrows);H-I-Emiliania huxleyi with etching and/or incomplete shield formation(arrows).All samples from the on-land record of Aravonitsa Plateau,location L11.
At the present day E.huxleyi type A,characterized by moderately elevated distal shield,robust distal shield elements and a central area covered by curved rod-like elements(Young et al.,2003),is abundant in the Mediterranean Sea.It dominates particularly the winter coccolithophore assemblages of the Aegean Sea.Overall,high cell densities are observed in winter and early spring during phases of highernutrient,well-mixed waters(Jan.:34×103cells/dm3,March:23×103cells/dm3),whereas densities are lower in the summer-early autumn oligotrophic and highly stratified water column(Aug.-Sept.:2×103cells/dm3;Dimiza et al.,2008;Triantaphyllou et al.,2010a).The analysis of sediment trap samples has shown that E.huxleyi largely dominates the total coccolith flux in the modern Aegean Sea and the eastern Mediterranean(Malinverno et al.,2009,2014;Triantaphyllou et al.,2004;Ziveri et al.,2000).
This study is based on the analysis of the coccolithophore assemblages occurring in marine sediments belonging to the NN21a and NN21b biozones that have been recovered from theeasternMediterraneanandfrommainlandGreece.Particular attention has been given to the variations in the pattern of E.huxleyi assemblages and their use both as abiostratigraphic tool and as a means to evaluate the palaeoenvironmental and palaeoclimatic conditions prevailing in these biozones.
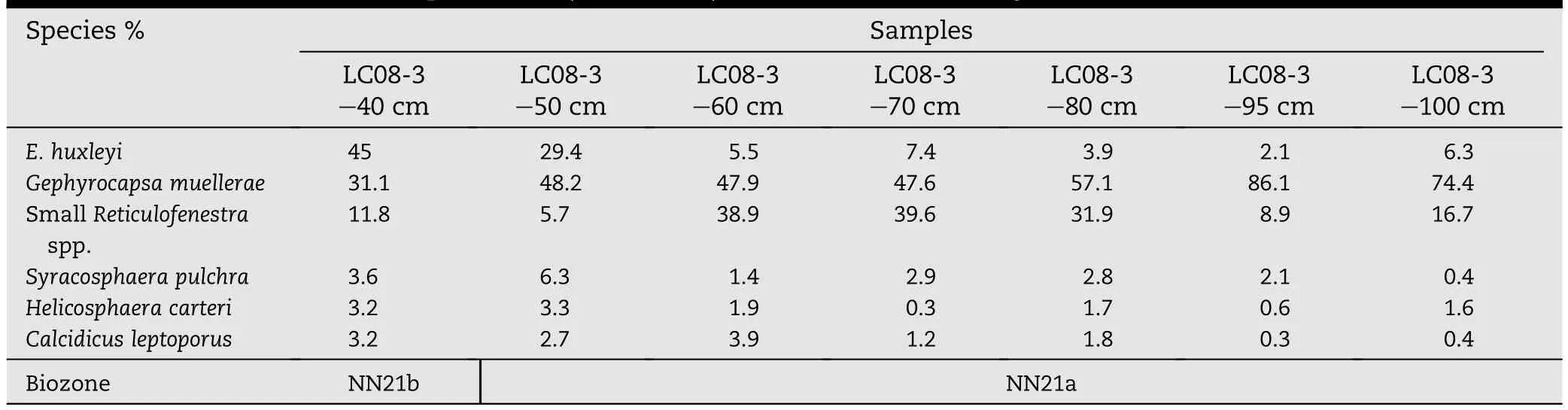
Table 2-Piston core LC08,sample LC08-3(40-100 cm):Quantitative SEM analysis of 300 coccoliths.
2.Samples and methods
The locations of the sample sites are shown in Fig.1.The single mainland site is on the Aravonitsa Plateau,northern Peloponnese(location L11,Palyvos et al.,2010).There are deep sea gravity cores from the Libyan Sea (ADE3-23:Triantaphyllou et al.,2010a),the southeastern Aegean Sea(NS-14:Triantaphyllou et al.,2009),and the southern Crete margin(HCMR2-22:Ioakim et al.,2009;Katsouras et al.,2010)as well as a long pistoncore from the Pantelleria Trough(LC08:Anastasakis and Pe-Piper,2006).
Samples of fine-grained marine sediments were examined by scanning electron and light microscopy for their content of calcareous nannoplankton using standard methods with counts of 300 coccolith specimens(Negri and Giunta,2001;Perch-Nielsen,1985;Thierstein et al.,1977).The definition of the E.huxleyi NN21a biozone is based on the first appearance of specimens of the species,that is the midpoint of the slope of the initial increase of the species in counts of 300 coccoliths(FAD of E.huxleyi;Rio et al.,1990;Thierstein et al.,1977),whereas the E.huxleyi Acme Zone(MNN21b)is defined as the interval where the frequency of E.huxleyi in the coccolith population exceeds 40%level(Castradori,1993;Rio et al.,1990).
3.Results
3.1.Terrestrial records

Fig.3-A-NN21a biozone assemblage,sample LC08-3-100 cm;B-E.huxleyi distal,sample LC08-3-100 cm;C-E.huxleyi distal,sample LC08-3-60 cm;D-E.huxleyi proximal,sample LC08-3-40 cm;E-Gephyrocapsa muellerae coccosphere,sample LC08-3-60 cm;F-Helicosphaera carteri proximal view,sample LC08-3-60 cm;G-H-Coccolithus pelagicus distal and proximal,sample LC08-3-60 cm;I-Helicosphaera carteri HOL(Syracolithus confusus),sample LC08-3-60 cm,from piston core LC08.
The sedimentary successions of fine-grained shallow marine deposits on the Aravonitsa Plateau are remnants preserved on a flat depositional geomorphic surface now at 500-520 m abovesea level.They werediscoveredduringthe investigation of the Pleistocene coastal uplift in the westernmost part of the Corinth Gulf(Palyvos et al.,2010).Two samples of these finegrained terrigenous sediments (location L11,samples W146A,W146B;Palyvos et al.,2010)yielded sparse nannoplankton assemblages.These contained abundant small Gephyrocapsa spp.(Table 1)including Gephyrocapsa protohuxleyi.The G.protohuxleyi specimens possess a bridge or the remains of the bases of a bridge(Fig.2A-E).It is also clear in each of these specimens that the distal shield elements are more or less tube-like with parallel sides,and there is some evidence in each instance that these had expanded ends(hammer heads)although most elements have lost at least part of their ends.In addition,small coccoliths resembling E.huxleyi occur very occasionally,and these bear platy distal shield elements in contact for much of their length and do not show any trace of hammer-heads.They have been identified as small specimens of Reticulofenestra spp.which have undergone etching resulting in separation of the ends of the elements(Fig.2F-G).Exhaustive microscopic analysis revealed the presence of rare(approximately 1%)true representatives of E.huxleyi with etching and/or incomplete shield formation(Table 1,Fig.2H-I).
The very low abundance of E.huxleyi in these samples could suggest correlation with the basal part of NN21a within the cold MIS 8 stage.Apparently,the lowest occurrence of E.huxleyi(at 270 ka)in the stratigraphic record of the eastern Mediterranean falls within the cold MIS 8,a phase of low sea levels based on sea-level curves(e.g.,Shackleton and Pisias,1985;Siddall et al.,2007).However,during this phase,the Corinth Gulf developed as a lake,isolated from the sea by a sill(e.g.,Palyvos et al.,2010).Depending on the elevation of the sill at the time,the lowest occurrence of E.huxleyi in the Corinth Gulf isthereforeexpectedto be moreor less closeto the MIS 7e highstand(i.e.a few ka before 240 ka),because only then sea water had the chance to enter the gulf(Palyvos et al.,2010).Therefore these marine deposits from the Aravonitsa Plateau(location L11)most probably belong to the warm MIS 7e stage at approx.240 ka,otherwise E.huxleyi would have been more abundant.
This is one of the very rare records of this marine biozone preserved in Greece.Another suggested terrestrial occurrence of this marine biozone has been described from a borehole drilled in the shallow valley of Livadi separating Kefallonia from its western peninsula,Paliki,during the search for Homer's Ithaca(Underhill,2006,2008).Very well preserved specimens of E.huxleyi have been recorded in the top 40 m of sediment of this borehole.It is suggested that these very young deposits may not represent in situ sedimentation,but can be considered as consistent with the impact of a catastrophic landslide on a marine channel.This would have ejected a large volume of water out of the channel and saturated the in fill material.This interpretation has been favored,because it was thought that E.huxleyi could not have reached this location earlier than about 6000 years ago,when rising global sea levels penetrated the shallow gulf of Livadi for the first time(Underhill,2008).
3.2.Marine cores

Fig.4-a-Stratigraphy,MIS stages and biozonation with core ADE3-23 from the Libyan Sea.The distribution of E.huxleyi and the top of G.muellerae Acme Zone are shown(modified from Triantaphyllou et al.,2010a);b-Stratigraphy,AMS ages and biozonal assignment of core HCMR2-22 from the southern Crete margin;c-The distribution of E.huxleyi and sea surface temperature(SST)pattern within the MIS 1 stage core NS-14 from southeastern Aegean Sea(modified from Triantaphyllou et al.,2009).S1a=The lower part of sapropel S1 layer;S1b=The upper part of sapropel S1 layer;SMH=Sapropel MidHolocene layer.
In the present study,a very clear transition between the NN21a and NN21b biozones is documented from the lower part of the long piston core LC08 in the Pantelleria Trough,Sicily Channel.This part of the core is dominated by the presence of a thick volcaniclastic interval deposited from a large gravitational flow during the Green Tuff eruption(Anastasakis and Pe-Piper,2006).The lower hemipelagic marls,immediately below the base of the volcaniclastic interval,have been examined for their nannofossil content.E.huxleyi is present in amounts of below 10%in the lower parts of this interval,and this rapidly increases upwards to 40%and more(Table 2,Fig.3).The presence of G.muellerae has been constantly recorded as more than 40%.A sample of marl from the hemipelagic interval yielded an accelerator mass spectrometry conventional radiocarbon age of 41190±1090 a BP(Beta 198833)with a calibrated radiocarbon age of 43-47±2 ka(Anastasakis and Pe-Piper,2006).Although radiocarbon dates older than 40-45 ka are questionable,the calcareous nannofossil assemblages confirm the biostratigraphic correlation of this interval(sample LC08-3-40 cm)with biozone NN21b,providing a unique chance to calibrate the base of this zone in the Mediterranean with an AMS radiocarbon age.
A case study involving the definition of NN21a/NN21b boundary has been documented in the gravity core ADE3-23,located SW of Crete(34°45.000′N,21°52.800′E),at 2459 m water depth.The core recovered 3.49 m of gray hemipelagic mud and silty mud interlayered with three distinct dark gray sapropelic layers,i.e.,S6,S5 and S1,and a hiatus of approximately 35 ka between ~83 ka and ~118 ka(Triantaphyllou et al.,2010a).E.huxleyi displays values well below 20%(Fig.4a)during the cold MIS 6 and MIS 4 stages,although during the latter,values are comparably higher.It just exceeds 10%during the warm MIS 5(sapropel S5 depositional interval).Apparently the lower part of the core ADE3-23 corresponds to the NN21a biozone.The increase in abundance of E.huxleyi,although never reaching 20%as previously reported(e.g.,Castradori,1993;Corselli et al.,2002;Violanti et al.,1991),suggests higher fertility in the surface waters in the middle part of S5,corresponding to the timing of relaxation in the northward penetration of the African monsoon already recorded within S5 in the southeastern Mediterranean area(Rohling et al.,2002).The definition of the NN21a/NN21b boundary is marked by the increase of E.huxleyi within MIS 3 at 50ka.Thiseventis shortlyfollowedin core ADE3-23 by the top of the Gephyrocapsa muellerae Acme Zone,characterized by a dramatic and continuous reduction in G.muellerae,which points to an age of 45.7 ka(Triantaphyllou et al.,2010a).A further increase of E.huxleyi is recorded within MIS 1(Fig.4a).

Fig.5-A-B-E.huxleyi Acme Zone assemblage,sample 67-68 cm;C-Gephyrocapsa muellerae,G.muellerae Acme Zone,samples 145-146 cm,and 173-174 cm;D-H-G.muellerae Acme Zone assemblage,samples 118-119 cm,145-146 cm,and 173-174 cm;I-Small Reticulofenestra spp.,sample 173-174 cm.Samples from gravity core HCMR2-22.
The top of the G.muellerae Acme Zone(45.7 ka BP)has also been recognized in the lower part of one additional sediment succession from gravity core HCMR2-22(34°33.968′N,24°53.770′E,water depth 2211 m,length 175 cm)from the southern Crete margin(Ioakim et al.,2009;Katsouras et al.,2010).Based on calcareous nannofossil biostratigraphy,most of this sedimentary succession has been proved to span the last approximately 50 ka and can be correlated with the E.huxleyi Acme Zone NN21b(Figs.4b and 5).
Very detailed information for the distribution of E.huxleyi during theLate Glacial-Holoceneintervalin thewarmMIS 1 is provided from a high-resolution record in the southeastern Aegean Sea(gravity core NS14;Triantaphyllou et al.,2009).The high surface nutrient indicator E.huxleyi(Young,1994)is the most abundant placolith within the coccolithophore assemblages from~13 to 10.6 ka BP(Fig.4c;Triantaphyllou et al.,2009).This points to cool surface-water conditions,as E.huxleyi is a species that prevails in the Aegean surface waters mainly in winter(Dimiza et al.,2008)and displays more heavily calcified E.huxleyi morphotypes(Triantaphyllou et al.,2010b).Following this interval,E.huxleyi decreases during the warm and wet phase during the deposition of sapropel S1.There is a prominent reduction starting at 8.5 ka BP with minimum values of 25%at the base of the upper part of sapropel S1(S1b),implying low surface productivity and strong stratification of the water column(Triantaphyllou et al.,2009).E.huxleyi increases again from 7.3 ka BP presenting three main negative shifts at~6.8,6.0 and 4.8 ka BP within the Sapropel Mid Holocene layer(SMH;Triantaphyllou et al.,2009,2014)associated with fluctuating sea-surface temperatures.(Fig.4c;Triantaphyllou et al.,2009).
4.Conclusions
The sediment records reveal variations in the E.huxleyi assemblages during marine isotope stages MIS 1-8.These appear to be linked with climatic variability.
In particular,based on E.huxleyi assemblages:
1)The E.huxleyi assemblage from the lowermost part of biozone NN21a is described from the Aravonitsa Plateau in the northern Peloponnese and most probably represents the warm phase MIS 7e at approx.240 ka.This is the first marine representative of this biozone onshore.The E.huxleyi coccoliths in the Kefallonia deposits are considered to be redeposited material.
2)The lower part of the long piston core LC08 from the Pantelleria Trough,Sicily Channel,displaysthe transition from biozone NN21a to NN21b.A radiocarbon(AMS)age of 41190±1090 a BP from this core provides a temporal calibration for the basal part of biozone NN21b in the eastern Mediterranean.
3)E.huxleyi displays values well below 20%of the total nannofossil assemblage during the cold stages MIS 6 and MIS 4 in the marine core ADE3-23 from the Libyan Sea,although during the latter stage,values are comparably higher.They exceed slightly 10%during the middle part of sapropel S5,within the warm MIS 5.
4)The biozone NN21a/NN21b boundary is marked by an abrupt increase of E.huxleyi within the relatively warm stage MIS 3 at 50 ka.Overlying this boundary in core ADE3-23 from the eastern Mediterranean is the top of the Gephyrocapsa muellerae Acme Zone at 45.7 ka;this has also been recognized in the lower part of the succession in core HCMR2-22 from the southern Crete margin.
5)A further increase and considerable fluctuations in E.huxleyi are recognized within warm stage MIS 1 in the core NS-14 from the southeastern Aegean Sea.This is thought to reflect continuous climate variability during this stage in the Late Glacial-Holocene.
Acknowledgements
G.Anastasakis is kindly thanked for providing the study material of piston core LC08.The manuscript benefited by the criticism of Christopher Jeans and two anonymous reviewers.
 Journal of Palaeogeography2015年4期
Journal of Palaeogeography2015年4期
- Journal of Palaeogeography的其它文章
- Sedimentary facies and lithologic characters as main factors controlling hydrocarbon accumulations and their critical conditions
- Correction of two Upper Paleozoic stratigraphic units in the Tianshan Mountains region,Xinjiang Uygur Autonomous Region and implications on the Late Paleozoic evolution of Tianshan tectonic complex,Northwest China
- Lithofacies palaeogeography of the Carboniferous and Permian in the Qinshui Basin, Shanxi Province, China
- Modeling the Middle Jurassic ocean circulation
- Correction of two Upper Paleozoic stratigraphic units in the Tianshan Mountains region, Xinjiang Uygur Autonomous Region and implications on the Late Paleozoic evolution of Tianshan tectonic complex,Northwest China
- End-Triassic nonmarine biotic events