
Neuregulin 1 isoforms could be an effective therapeutic candidate to promote peripheral nerve regeneration
2014-06-01 09:42:34GiovannaGambarotta,GiuliaRonchi,StefanoGeuna等
中國神經(jīng)再生研究(英文版) 2014年12期
Neuregulin 1 isoforms could be an effective therapeutic candidate to promote peripheral nerve regeneration
Traumatic injuries of peripheral nerves represent common casualties and their social impact is considerably high. Although peripheral nerves retain a good regeneration potential, the clinical outcome after nerve lesion is far from being satisfactory and functional recovery is almost never complete, especially in the case of large nerve defects, that result in loss or diminished sensitivity and/or motor activity of the innervated target organs. Therefore, to improve the outcome after nerve damage, or in peripheral neuropathies, there is a need for further research in nerve repair and regeneration to identify factors that promote axonal regrowth, remyelination and target reinnervation.
Among the different factors involved in these processes (Taveggia et al., 2010; Pereira et al., 2012), stands out neuregulin 1 (NRG1), a factor which plays a role both in the myelination occurring during development (Lemke, 2006) and in the response to peripheral nerve injury (Syed and Kim, 2010; Fricker and Bennett, 2011).
NRG1 is a pleiotropic factor characterized by the existence of numerous isoforms arising from alternative splicing of exons that confer to the protein with deeply different characteristics (Falls, 2003; Mei and Xiong, 2008).
NRG1 can be produced as a secreted or as a transmembrane protein ready to interact with its receptor, or as a transmembrane pro-protein that needs a proteolytic cleavage to release a soluble fragment or to protrude its receptor binding domain in the extracellular environment (Figure 1). According to its structure, NRG1 signals in a paracrine, autocrine or juxtacrine manner; moreover, juxtacrine interactions can signal both in a forward and reverse manner due to the production of a fragment containing the intracellular domain (ICD, Figure 1B) that can translocate into the nucleus and in fl uence gene transcription (Bao et al., 2003; Bao et al., 2004; Chen et al., 2010). NRG1 interacts directly with two of the four members of the tyrosine kinase receptor family ErbB: ErbB4, that signals as homo or heterodimer, and ErbB3, that forms a heterodimer with ErbB2.
In the peripheral nervous system, NRG1 soluble isoforms are mainly released by Schwann cells, while transmembrane isoforms are mainly expressed by the axon and both interact with the heterodimer receptor ErbB2-ErbB3, generally expressed by Schwann cells. NRG1 plays an important role both in the myelination occurring during development and in the different phases occurring after injury in the peripheral nerve: axon degeneration, axon regrowth, remyelination and target reinnervation (Taveggia et al., 2010; Fricker and Bennett, 2011; Pereira et al., 2012; Salzer, 2012; Gambarotta et al., 2013; Heermann and Schwab, 2013).
These processes respond to different cues, as can be inferred from the analysis of transgenic mice models summarized in Figure 2. The difference between the myelination process occurring during development and the regeneration process occurring after nerve injury is underlined by the fact that soluble NRG1 isoforms play an important role after nerve injury, while their lack seems irrelevant during development.
Membrane bound NRG1 determinates the myelination fate during development
During development, the absence of soluble NRG1 in Schwann cells does not affect myelination (Stassart et al., 2013) and, accordingly, soluble NRG1 over-expression in motoneurons and dorsal root ganglia (DRG) neurons does not in fl uence myelination (Michailov et al., 2004).
Conversely, axonal transmembrane NRG1 expression level determines the myelination fate of axons and the thickness of the myelin sheath: animals lacking axonal transmembrane NRG1 show hypomyelination (Michailov et al., 2004; Taveggia et al., 2005), while its over-expression causes hypermyelination (Michailov et al., 2004) and conversion of normally non-myelinated neurons to myelinated neurons (Taveggia et al., 2005).
Soluble and membrane bound NRG1 play different roles after peripheral nerve injury
Animals lacking soluble NRG1 in Schwann cells display peripheral nerve regeneration severely impaired (Stassart et al., 2013). Accordingly, soluble NRG1 over-expression in motoneurons and dorsal root ganglion neurons improves remyelination after injury (Stassart et al., 2013).
Immediately after injury, we (unpublished results) and others (Stassart et al., 2013) observed that the soluble NRG1 transcript is strongly upregulated in the distal and proximal nerve. Because RNA extracted from the nerve belongs mainly to Schwann cells, this observation suggests that Schwann cells, following nerve injury, produce high amounts of soluble NRG1 that could stimulate, in an autocrine manner, Schwann cell survival and, likely, migration of macrophages that remove myelin debris in the early phases of Wallerian degeneration to allow remyelination (Fricker and Bennett, 2011).
The soluble NRG1 upregulation observed in Schwann cells immediately after nerve injury suggests that denervated Schwann cells require autocrine stimulation with soluble NRG1 for survival and that the peripheral nerve regeneration impairment observed in animals lacking Schwann cell derived soluble NRG1 is the indirect consequence of problems occurring during the early phases of axon degeneration and axon regrowth, not during the following phases of remyelination and target reinnervation.
Animals lacking axonal transmembrane NRG1 isoforms show an impaired rate of remyelination and functional recovery at early phases after nerve injury; at later stages, the myelination thickness is not strictly dependent on axonal NRG1 and it has been hypothesized a compensation effect mediated by other factors (Fricker et al., 2013). Accordingly, axonal transmembrane NRG1 over-expression improves peripheral nerve regeneration (Stassart et al., 2013).
Strategies to promote nerve regeneration
These observations suggest that soluble NRG1 plays a role during the early phases following nerve injury corresponding to axon degeneration and regrowth, while transmembrane NRG1 plays a role during later phases corresponding to the remyelination process. Therefore, soluble NRG1, already used in human trials for heart failure treatment, could be an effective therapeutic candidate to promote nerve regeneration. Accordingly, it has been already demonstrated that nerve regeneration is successfully promoted by subcutaneous NRG1 injection (Chen et al., 1998; Yildiz et al., 2011), by NRG1 released by biomaterials (Mohanna et al., 2003; Cai et al., 2004; Mohanna et al., 2005) or by adenovirus encoded NRG1 (Joung et al., 2010). Moreover, it has been suggested that NRG1 is released by the degenerating muscle successfully used to fi ll a non-nervous conduit graft consisting of a vein to bridge the proximal and the distal stumps after substance loss (Nicolino et al., 2003).
However, we think that treatment with recombinant soluble NRG1 should be carried out in a well-defined time window, during early phases following nerve injury, to improve survival, migration and redifferentiation of Schwann cells, in synergy with endogenous NRG1 released by Schwann cells immediately after injury, that in cases of severe damage may not be suf fi cient.
Furthermore, NRG1 treatment should be fi nely regulated, because it has been demonstrated in vitro that different NRG1 isoforms have different pro-myelinating activities and a too high concentration can inhibit myelination (Syed et al., 2010).
A second strategy to promote myelination could be the over-expression of recombinant transmembrane NRG1 in axons during later phases following nerve injury. However, to express transmembrane isoforms, the use of viral vectors would be necessary; to bypass this critical step, manipulation of the processing of endogenously expressed NRG1 could increase its pro-myelinating activity. Actually,transmembrane NRG1 can be cleaved by different metalloproteases, including the α secretase TACE (also known as ADAM17) and the β secretase BACE1, and other not yet identi fi ed proteases, that cleave the transmembrane NRG1 in the same stalk region, leaving the EGF-like domain exposed and C terminal domains that differ by a few amino acids (Figure 1).
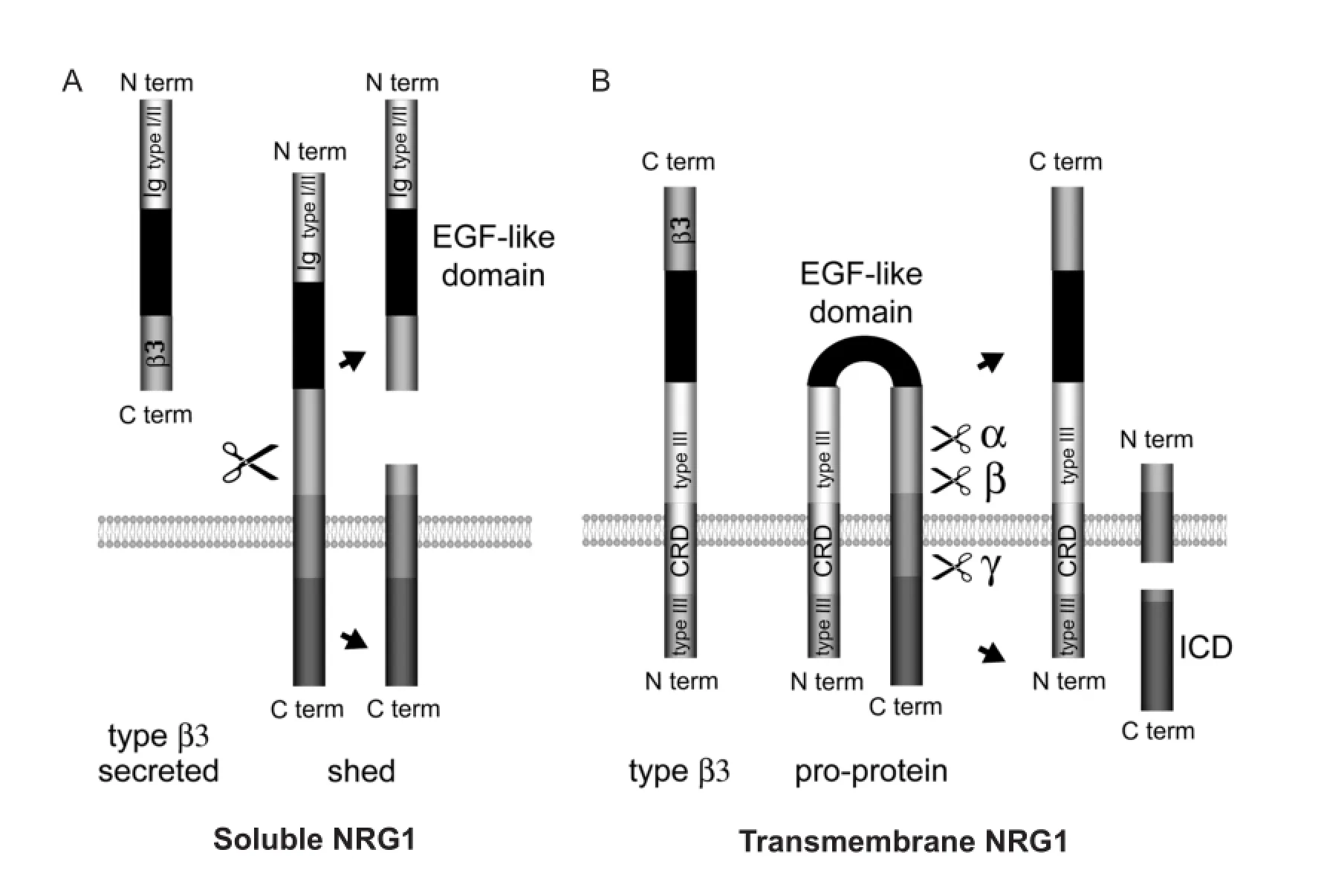
Figure 1 Structure of soluble or transmembrane neuregulin 1 (NRG1) isoforms.
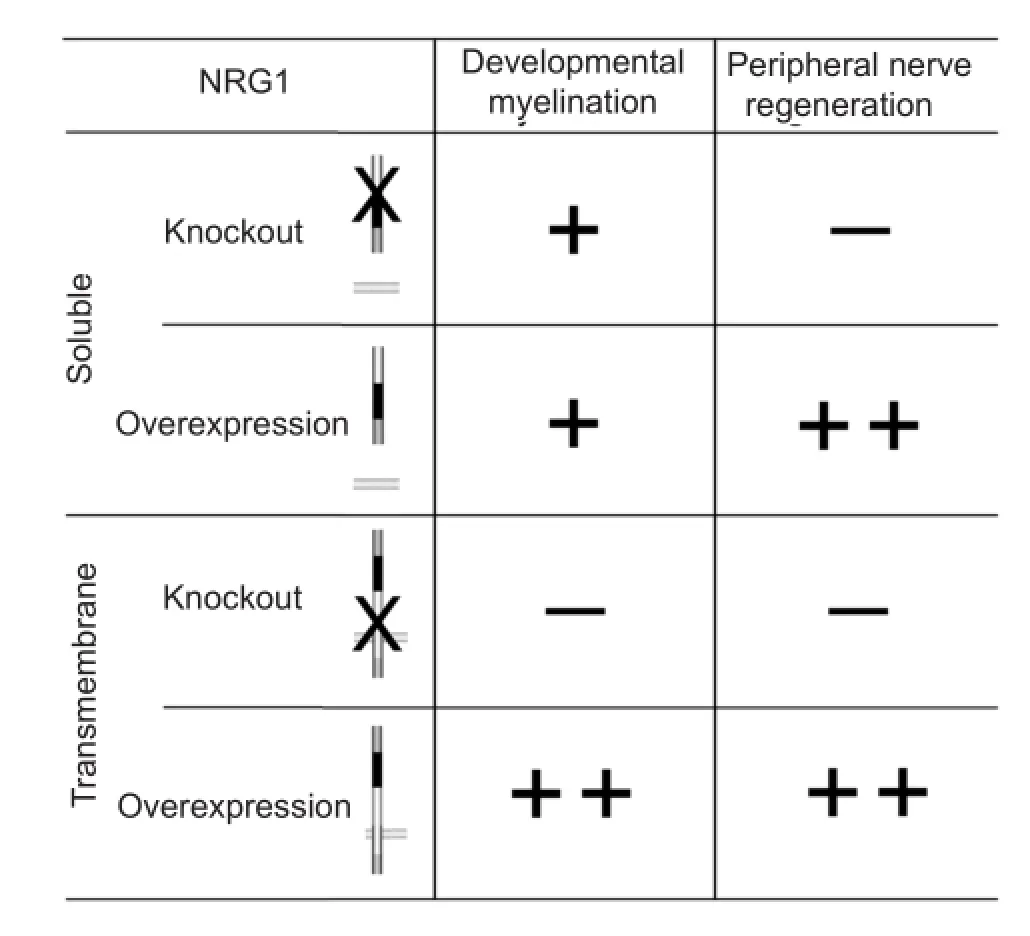
Figure 2 The role played by soluble and transmembrane neuregulin 1 (NRG1) isoforms in the myelination occurring during development and in the different phases occurring after nerve injury (axon degeneration, axon regeneration, remyelination and target reinnervation) as inferred from transgenic and conditional knockout mice.
The effect on myelination of these proteases seems to be opposite: the β secretase BACE1 cleavage activates the pro-myelinating activityof NRG1, as shown in BACE1 knockout mice characterized by an hypo-myelination phenotype (Willem et al., 2006; Hu et al., 2008) and in transgenic mice over-expressing a recombinant NRG1 mimicking the BACE1 cleavage, characterized by an hyper-myelinated phenotype (Velanac et al., 2012). It would be interesting to analyze the remyelination ef fi ciency in these mice, to understand if BACE1 plays a role only during developmental myelination or also during remyelination occurring after peripheral nerve injury and repair. However, a pro-myelinating strategy including the treatment with BACE1 stimulators, if any, would not be recommended, because BACE1 is a major drug target for Alzheimer’s disease: BACE1-mediated cleavage of amyloid precursor protein (APP) is the fi rst step in the generation of the pathogenic amyloid-β peptides and recent studies demonstrate a wide range of BACE1 physiological substrates and functions (Vassar et al., 2014).
Conversely, the α secretase TACE cleavage inhibits the pro-myelinating activity of NRG1 and its inactivation in motor neurons -obtained through conditional knockout mice-correlates with a hyper-myelination phenotype during development and in the adult (La Marca et al., 2011). No data concerning remyelination ef fi ciency following peripheral nerve injury in mice in which TACE is inactivated or inhibited by pharmacological treatments are available and it would be really useful to test if TACE inactivation promotes remyelination during peripheral nerve regeneration.
Different TACE inhibitors are already available and used in preclinical trials anti rheumatoid arthritis and anti breast cancer (DasGupta et al., 2009; Rego et al., 2014) and could be useful tools to promote remyelination.
Because regeneration is spontaneous, but often incomplete, the development of new strategies to promote peripheral nerve regeneration is a significant goal to achieve, and the pleiotropic NRG1 isoforms appear to be good candidates for therapeutic treatments.
Giovanna Gambarotta1,2, Giulia Ronchi1,3, Stefano Geuna1,2,3, Isabelle Perroteau1,2
1 Department of Clinical and Biological Sciences, University of Turin, Italy
2 Neuroscience Institute of Turin (NIT), University of Turin, Italy
3 Neuroscience Institute of the “Cavalieri Ottolenghi” Foundation (NICO), University of Turin, Italy
Bao J, Wolpowitz D, Role LW, Talmage DA (2003) Back signaling by the Nrg-1 intracellular domain. J Cell Biol 161:1133-1141.
Bao J, Lin H, Ouyang Y, Lei D, Osman A, Kim TW, Mei L, Dai P, Ohlemiller KK, Ambron RT (2004) Activity-dependent transcription regulation of PSD-95 by neuregulin-1 and Eos. Nat Neurosci 7:1250-1258.
Cai J, Peng X, Nelson KD, Eberhart R, Smith GM (2004) Synergistic improvements in cell and axonal migration across sciatic nerve lesion gaps using bioresorbable fi laments and heregulin-beta1. J Biomed Mater Res A 69:247-258.
Chen LE, Liu K, Seaber AV, Katragadda S, Kirk C, Urbaniak JR (1998) Recombinant human glial growth factor 2 (rhGGF2) improves functional recovery of crushed peripheral nerve (a double-blind study). Neurochem Int 33:341-351.
Chen Y, Hancock ML, Role LW, Talmage DA (2010) Intramembranous valine linked to schizophrenia is required for neuregulin 1 regulation of the morphological development of cortical neurons. J Neurosci 30:9199-9208.
DasGupta S, Murumkar PR, Giridhar R, Yadav MR (2009) Current perspective of TACE inhibitors: a review. Bioorg Med Chem 17:444-459.
Falls DL (2003) Neuregulins: functions, forms, and signaling strategies. Exp Cell Res 284:14-30.
Fricker FR, Bennett DL (2011) The role of neuregulin-1 in the response to nerve injury. Future Neurol 6:809-822.
Fricker FR, Antunes-Martins A, Galino J, Paramsothy R, La Russa F, Perkins J, Goldberg R, Brelstaff J, Zhu N, McMahon SB, Orengo C, Garratt AN, Birchmeier C, Bennett DL (2013) Axonal neuregulin 1 is a rate limiting but not essential factor for nerve remyelination. Brain 136:2279-2297.
Gambarotta G, Fregnan F, Gnavi S, Perroteau I (2013) Neuregulin 1 role in Schwann cell regulation and potential applications to promote peripheral nerve regeneration. Int Rev Neurobiol 108:223-256.
Heermann S, Schwab MH (2013) Molecular control of Schwann cell migration along peripheral axons: keep moving! Cell Adh Migr 7:18-22.
Hu X, He W, Diaconu C, Tang X, Kidd GJ, Macklin WB, Trapp BD, Yan R (2008) Genetic deletion of BACE1 in mice affects remyelination of sciatic nerves. FASEB J 22:2970-2980.
Joung I, Yoo M, Woo JH, Chang CY, Heo H, Kwon YK (2010) Secretion of EGF-like domain of heregulinbeta promotes axonal growth and functional recovery of injured sciatic nerve. Mol Cells 30:477-484.
La Marca R, Cerri F, Horiuchi K, Bachi A, Feltri ML, Wrabetz L, Blobel CP, Quattrini A, Salzer JL, Taveggia C (2011) TACE (ADAM17) inhibits Schwann cell myelination. Nat Neurosci 14:857-865.
Lemke G (2006) Neuregulin-1 and myelination. Sci STKE 2006:pe11.
Mei L, Xiong WC (2008) Neuregulin 1 in neural development, synaptic plasticity and schizophrenia. Nat Rev Neurosci 9:437-452.
Michailov GV, Sereda MW, Brinkmann BG, Fischer TM, Haug B, Birchmeier C, Role L, Lai C, Schwab MH, Nave KA (2004) Axonal neuregulin-1 regulates myelin sheath thickness. Science 304:700-703.
Mohanna PN, Terenghi G, Wiberg M (2005) Composite PHB-GGF conduit for long nerve gap repair: a long-term evaluation. Scand J Plast Reconstr Surg Hand Surg 39:129-137.
Mohanna PN, Young RC, Wiberg M, Terenghi G (2003) A composite poly-hydroxybutyrate-glial growth factor conduit for long nerve gap repairs. J Anat 203:553-565.
Nicolino S, Raimondo S, Tos P, Battiston B, Fornaro M, Geuna S, Perroteau I (2003) Expression of alpha2a-2b neuregulin-1 is associated with early peripheral nerve repair along muscle-enriched tubes. Neuroreport 14:1541-1545.
Pereira JA, Lebrun-Julien F, Suter U (2012) Molecular mechanisms regulating myelination in the peripheral nervous system. Trends Neurosci 35:123-134.
Rego SL, Helms RS, Dreau D (2014) Tumor necrosis factor-alpha-converting enzyme activities and tumor-associated macrophages in breast cancer. Immunol Res 58:87-100.
Salzer JL (2012) Axonal regulation of Schwann cell ensheathment and myelination. J Peripher Nerv Syst 17 Suppl 3:14-19.
Stassart RM, Fledrich R, Velanac V, Brinkmann BG, Schwab MH, Meijer D, Sereda MW, Nave KA (2013) A role for Schwann cell-derived neuregulin-1 in remyelination. Nat Neurosci 16:48-54.
Syed N, Kim HA (2010) Soluble neuregulin and Schwann cell myelination: a therapeutic potential for improving remyelination of adult axons. Mol Cell Pharmacol 2:161-167.
Syed N, Reddy K, Yang DP, Taveggia C, Salzer JL, Maurel P, Kim HA (2010) Soluble neuregulin-1 has bifunctional, concentration-dependent effects on Schwann cell myelination. J Neurosci 30:6122-6131.
Taveggia C, Feltri ML, Wrabetz L (2010) Signals to promote myelin formation and repair. Nat Rev Neurol 6:276-287.
Taveggia C, Zanazzi G, Petrylak A, Yano H, Rosenbluth J, Einheber S, Xu X, Esper RM, Loeb JA, Shrager P, Chao MV, Falls DL, Role L, Salzer JL (2005) Neuregulin-1 type III determines the ensheathment fate of axons. Neuron 47:681-694.
Vassar R, Kuhn PH, Haass C, Kennedy ME, Rajendran L, Wong PC, Lichtenthaler SF (2014) Function, therapeutic potential and cell biology of BACE proteases: current status and future prospects. J Neurochem 130:4-28.
Velanac V, Unterbarnscheidt T, Hinrichs W, Gummert MN, Fischer TM, Rossner MJ, Trimarco A, Brivio V, Taveggia C, Willem M, Haass C, Mobius W, Nave KA, Schwab MH (2012) Bace1 processing of NRG1 type III produces a myelin-inducing signal but is not essential for the stimulation of myelination. Glia 60:203-217.
Willem M, Garratt AN, Novak B, Citron M, Kaufmann S, Rittger A, DeStrooper B, Saftig P, Birchmeier C, Haass C (2006) Control of peripheral nerve myelination by the beta-secretase BACE1. Science 314:664-666.
Yildiz M, Karlidag T, Yalcin S, Ozogul C, Keles E, Alpay HC, Yanilmaz M (2011) Efficacy of glial growth factor and nerve growth factor on the recovery of traumatic facial paralysis. Eur Arch Otorhinolaryngol 268:1127-1133.
Giovanna Gambarotta, Dipartimento di Scienze Cliniche e Biologiche, Università di Torino, Ospedale San Luigi, Regione Gonzole 10, 10043 - Orbassano (TO), Italy, giovanna.gambarotta@unito.it. Acknowledgments: We apologize for any omissions in citing relevant publications. The research leading to this paper has received funding from the European Community’s Seventh Framework Programme (FP7-HEALTH-2011) under grant agreement No. 278612 (BIOHYBRID), from MIUR and from Compagnia di San Paolo (MOVAG).
10.4103/1673-5374.135324 http://www.nrronline.org/
Accepted: 2014-05-16
Gambarotta G, Ronchi G, Geuna S, Perroteau I. Neuregulin 1 isoforms could be an effective therapeutic candidate to promote peripheral nerve regeneration. Neural Regen Res. 2014;9(12):1183-1185.

- 中國神經(jīng)再生研究(英文版)的其它文章
- 1111
- Overexpression of microRNA-124 promotes the neuronal differentiation of bone marrow-derived mesenchymal stem cells
- Assessment on self-care, mobility and social function of children with spina bi fi da in Turkey
- Recovery of the corticospinal tracts injured by subfalcine herniation: a diffusion tensor tractography study
- Diffuse axonal injury after traumatic cerebral microbleeds: an evaluation of imaging techniques
- Autophagy: a double-edged sword for neuronal survival after cerebral ischemia