
SURVIVAL AND CARBOHYDRATE STORAGE IN TWO TOLERANT PLANT SPECIES EXPOSED TO PROLONGED FLOODING IN THE THREE GORGES RESERVOIR REGION
2013-07-24 11:29:38YEXiaoQiandZENGBo
水生生物學(xué)報 2013年3期
YE Xiao-Qi and ZENG Bo
(1.Institute of Subtropical Forestry, Chinese Academy of Forestry, Fuyang 311400, China; 2.Key Laboratory of Eco-environments in Three Gorges Reservoir Region (Ministry of Education), Chongqing Key Laboratory of Plant Ecology and Resources Research in
Three Gorges Reservoir Region, School of Life Science, Southwest University, Chongqing 400715, China)
Annual summer flooding often occurs in Yangtze River in the Three Gorges Reservoir (Yangtze River,China) region.Such natural flooding is characterized by temporary but high water table rise.Arundinella anomalaandSalix variegataare two of the flood-tolerant species growing in riverside.Our previous results showed these two species could survival at least three months complete submergence[1].The contrasting short natural river flooding regimes and high survival rate in artificially prolonged flooding suggest these species have a capable of resisting unexpected flooding stress.
When terrestrial plants are flooded, one of the most remarkable stresses is reduced gas exchange between plants and the environments due to much slower gas diffusion rates in water than in air[2,3].Shortage of CO2and O2will directly influence the carbon budget of the whole plants at this site.Upon flooding, carbon availability becomes one of the critical factors for plant survival.Photosynthetic assimilation rate can be either decreased in mild waterlogging conditions[4,5]or almost inhibited in submergence conditions[6].Even though potential of underwater photosynthesis has been shown in some riparian species, the realized carbon assimilation performance is limited by the low light and low carbon dioxide concentration in water[7].Carbohydrate utilization efficiency is further lowered by shortage of oxygen.Some studies reported that sugar oxidation by glycolysis in anoxia condition yields only two mol ATP per glucose,compare to thirty-six mol ATP in aerobic conditions[8],which suggests plants have to accelerate sugar utilization if the same amount ATP is demanded.Adapatations to reduce energy requirements and distribute energy to essential processes are highly valued in plants anoxia tolerance[9].It is commonly recognized that flood tolerance can be mostly explained by oxygen deficiency tolerance[10].The capacity to mobilize carbohydrate pool has been implicated in the mechanisms of oxygen deficiency tolerance in some wetland plants[11,12].Hence we hypothesize that plants which can survive during long-term flooding have to tolarate carbon starvation.One of the mechanisms on long-term flooding tolerance of species is the capacity of carbon storage mobilization.More generally, carbon starvation was considered as the key to reed decline in eutrophic lakes[13].However, few studies focus on carbohydrate metabolism in relation to flood tolerance under more natural conditions[14], with exception of rice plants, anyhow, with a very limited submergence tolerance[15,16].
Up to now, quite a lot of plant species have been reported to be flood-tolerant in the Three Gorges Reservoir region[1,17—19].However, little is known how survival in prolonged flooding couples with carbohydrate concentration in these species.In our previous work, simulated flooding decreased soluble sugar concentration and starch concentration ofS.variegataplants within 60 days treatments; complete submergence caused larger reduction of sugar concentration and starch concentration than waterloging[20].Further research showed thatS.variegatacould survive to 180 days of complete submergence;A.anomalaplants could also keep high photosynthetic capacity after 60 days of flooding treatments[1].How carbohydrate concentration in these two tolerant species changes with prolonged flooding and how it is related to survive rate need further investigation.
In this paper,A.anomalaandS.variegataplants were exposed to simulated and prolonged flooding conditions with three water levels.Survival rate after long-term flooding was examined and carbohydrate dynamics during the whole flooding period was characterized.We aimed to find out whether carbohydrate pool in these two species can be mobilized and what the possible relationship between carbohydrate level and survival rate of long-term flooding conditions.
1 Materials and Methods
1.1 Plant materials and growth conditions
In May 2005, clones ofA.anomalaand one-year oldS.variegataseedlings were collected from riverside of Jialing River, one of the largest branches of Yangtze River.To homogenise the initial plants size, single rametsA.anomalawith shoot length of 10 to 15 cm and seedlings ofS.variegatawith shoot height of 5—7 cm were selected and grown in plastic pots (15 cm tall, 20 cm diameter) filled with a 1∶4 (v∶v) of mixture of humus and loam.All the plants were placed inside in the ecology experimental garden in Southwest China University(Chongqing, China).The seedlings were growing under ambient climate conditions with sufficient tapwater supply.
1.2 Treatments
Treatments started in the greenhouse at the end of January 2006, with the same experiment design for the both species as the following: Before initiation of the experiments, 8 plants from the stock plants were harvested for biomass and carbohydrate analysis.The remaining plants were randomly allocated to three groups with twenty-eight plants in each group.One group plants were put in a concrete water basin (2.5 m × 2.0 m × 2.5 m) filled with tap water.The water depth was kept 2 cm above soil surface and only the belowground of the plants was flooded (waterlogging), a second group plants were completely submerged in an identical basin, with the water depth remaining at 2 m (submergence).The third group plants were left growing in well drained soils but well watered (controlled plants, CK).Seven plants from each group were harvested after 40, 90, 120 and 180 days treatments.The water in the basins was replaced once a week; for complete submerged plants, exposure to air by shoots was avoided by keeping the stable water depth.
1.3 Survival rate determination
At each harvesting time, for survival rate estimation,12 plants from each of the two flooding treatments (waterlogging and submergence) were returned to the same normal conditions as control plants.After two months of recovery, the number of living plants and dead plants were recorded.Plants without initiation of buds and green leaves were considered dead.The survival rate was calculated as the ratio of the number of living plants to the total number of 12 plants in each treatment.
1.4 Carbohydrate extraction and determination
For soluble sugar analysis, additional seven plants from each treatment were harvested at day 40, 90, 120 and 180.The plants were washed under tap water and divided into leaves, stems and roots, freeze-dried for 48h and ground to fine powder.About 30 mg powder from each sample was extracted in boiling 80% ethanol for 30min for three times.All the extracts from each sample were pooled together and final volume was made 25 mL.Soluble sugar concentration in each extract was determined with DNS (3, 5-dinitrosalicylic acid) at 540 nm using a spectrometer (UV-2550, SHIMADZU), according to the method by Miller[21].For starch analysis, the residues leaved from soluble extraction were hydrolyzed using 4 mL demi and 2 mL 8 M HCl (1h, 100℃), ac-cording to van Eck,et al.[22].The glucose released from starch were measured with the same method above.
1.5 Data analysis
All the statistical analysis was conducted in SPSS11.0.The average soluble sugar concentration and starch concentration in stem, leaf and root (forS.variegata,fine root and coarse root) were calculated.Two-way ANOVA was used to test the two fixed factor flooding water depth treatments (Control, waterlogging and submergence) and flooding duration effects (40d, 90d, 120d and 180d) on soluble sugar and starch concentration.When it was necessary, a Post Hoc multiple comparison with Least Significant Difference test (LSD,P= 0.05)was conducted.
2 Results
2.1 The effects of flooding on survival rate, biomass and carbohydrate concentration in A.anomala plants
All waterloggedA.anomalaplants survived 180 days treatment; about one quarter of the complete submerged plants survived 120 days treatment but all died when submerged for 180 days (Tab.1).
Both flooding duration and water depth had significant effects on plant biomass accumulation, except for stems (Tab.2).As flooding duration prolonged, dry mass of the roots, leaves and whole plants of the non-flooded plants and waterlogged plants kept increasing, while for submerged plants, dry mass of the roots remained almost stable (Fig.1a), and dry mass of the stems, leaves and whole plants kept decreasing (Fig.1).Waterlogging did not decrease biomass accumulation rate compared with non-flooded plants, except at 180d (Fig.1), while submergence decreased dry mass of all the three tissues significantly starting from day 90, compared with the control and waterlogging treatments (P< 0.05) (Fig.1).Significant effects of interaction between flooding duration and water depth on dry mass occurred for root, leaf and whole plant, but not for stem (Tab.2).
Flooding duration and water depth also imposed significant effects on carbohydrate concentration in the plants (Tab.3).Generally, waterlogging only led to slight reduction of soluble sugar and starch concentration (P>0.05, Fig.2).Significant reduction of soluble sugar and starch concentration occurred in submerged plants starting from day 90, compared with non-flooded and waterlogged treatments (P<0.05, Fig.2).In submerged plants,in stems and leaves, after a reduction at day 40, soluble sugar concentration remained relatively stable (P>0.05,Fig.2c & e), while in roots, soluble sugar concentration kept decreasing until to a much low level at the end of the treatment (P<0.05, Fig.2a).Significant effects of interaction between flooding duration and water depth occurred for both sugar and starch in all the tissues (Tab.3).

Tab.1 Survival rate (%) of A.anomala and S.variegata plants in non-flooding (control), waterlogging (2 cm water above soil surface) and submergence (2 m deep water) treatments.n = 12

Tab.2 F-values and significance of a two-way ANOVA of dry mass of A.anomala and S.variegata plants in 3 water depth treatments: non-flooding(control), waterlogging (2 cm water above soil surface) and submergence (2 m deep water) after 40 days, 90 days, 120 days and 180 days of treatments.
2.2 The effects of flooding on survival rate, dry mass accumulation and carbohydrate concentration in S.variegata plants
The floodedS.variegataplants survived all the treatments except for 180 days complete submergence (Tab.1).
There were great variation of dry mass of all tissues between flooding duration and water depth treatments in theS.variegataplants (Fig.3).Both flooding duration and water depth had significant effects on the dry mass accumulation (Tab.2), except for the dry mass of the fine roots, which were not sensitive to the water depth treatments (Fig.3b).Generally, as flooding duration prolonged, dry mass of the nonflooded plants and waterlogged plants kept increasing; for submerged plants, dry mass of the coarse roots, stems and whole plants remained almost stable (Fig.3b, c & e), while it decreased significantly for the leaf (P< 0.05, Fig.3d).There were significant effects of interaction between flooding duration and water depth on biomass of stems and leaves, but not on biomass of coarse roots, fine roots and whole plant (Tab.2).
Carbohydrate concentration dynamics in floodedS.variegataplants was very similar to those inA.anomalaplants (Fig.2 and Fig.4).With the exception of fine roots, waterlogging only decreased soluble sugar and starch concentration after 40 days in various plant parts and later recovered to the level in non-flooded plants(Fig.4).In fine roots, waterlogging caused a larger reduction of both soluble sugar and starch concentration(Fig.4a & b,P< 0.05).Complete submergence treatment reduced remarkably soluble sugar and starch concentration in all tissues.In roots and stems, the decrease of soluble sugar (Fig.4a, c & e) lasted during the whole flooding treatments while decrease of starch concentration (Fig.4b, d & f) seemed slowed down in the late period.After 90 days treatment, all the submerged plants shed their leaves; therefore no carbohydrate measurements were made (Fig.4g & h).There were significant effects of interaction between flooding duration and water depth for both sugar and starch in all tissues, except for starch in the roots (Tab.3).

Fig.1 Dry mass (g) in non-flooded (control), waterlogged (2 cm deep water above soil surface) and completely submerged (2 m deep water) A.anomala plants after 40, 90, 120 and 180 days of treatments.Points = means ± SE, n = 7
3 Discussion
Our results indicated high flooding tolerance in these two species, as shown in the high survival rate of long-term submergence (Tab.1).The high flood tolerance in these two species was comparable to some riparian species reported earlier[17—19,23].The two species could also accumulate a large amount of carbohydrate in non-flooded plants (Fig.2 and Fig.4).It might be that the large carbohydrate pool can be mobilized in the incoming annual summer flooding.
The results indicated that submergence had more detrimental effects on the two species than waterlogging,which did not lead to mortality in both plant, neither did it also reduce biomass accumulation (Fig.1 and Fig.3,P<0.05) and carbohydrate availability (Fig.2 and Fig.4),compared with non-flooded plants.In contrast, submergence reduced survival rate, hindered biomass accumulation and reduced carbohydrate concentration.This suggested that shoot exposure to air[24]and gas exchange of both CO2and O2were the dominate factors[7]for these two species in control of the responses to flooding, since submergence decreases photosynthetic carbon fixation[6]and leads to inefficient of carbohydrate consumption owing to insufficient oxygen supply[9], which were critical for growth and survival for the plants under water.The results also indicated significant interaction effects of flooding duration and water depth.This showed different dynamics of biomass and carbohydrate concentration in the different flooding treatments.Partly, this could be due to increasing air temperature from January to July.In another study, it has been found that higher water temperature led to faster carbohydrate consumption in submergedA.anomalaplants under controlled water temperature conditions (data not shown).The responses to air or water temperature may be different from nonflooded, waterlogged and submerged plants.In another study on the non-structural carbohydrate concentration in dynamics in roots ofS.variegataplants conducted from August to October[20], it was found that waterlogging and submergence decreased carbohydrate concentration to a greater extent compared to the results in our study;furthermore, their results[20]also showed a much larger effects of waterlogging and submergence duration effects on carbohydrate concentration.This may also due to the air temperature difference in the two studies.Since the flooding treatments in our study started from end January to end of July, which had a lower mean air temperature,much mild effects of flooding onS.variegatacould be due to lower air temperature.It has been shown that winter flooding had less negative effects on several plant species than winter flooding[22].
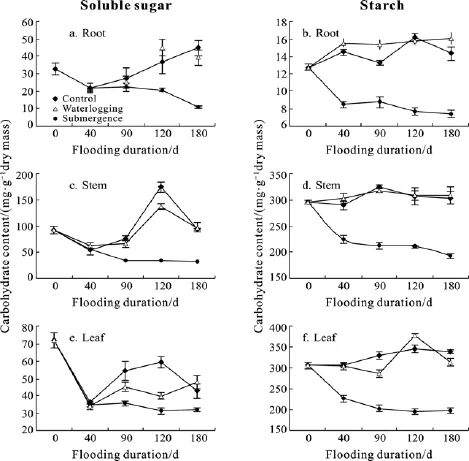
Fig.2 Soluble sugar and starch concentration (mg/g dry mass) in non-flooded (control), waterlogged (2 cm deep water above soil surface) and completely submerged (2 m deep water) A.anomala plants after 40, 90, 120 and 180 days of treatments.Points = means ± SE, n = 7
Low survival rate was coupled with large biomass loss and decreased of carbohydrate concentration with prolonged submergence.Biomass loss during the flooding period could be partly due to shedding of leaves (Fig.1c and Fig.2d) and partly due to consumption of carbohydrate (Fig.3 and Fig.4).Biomass loss was reported to be a characteristic of some flood-tolerant species[18,23].Probably, this loss of biomass is a consequence of replacement of old tissues by new tissues formation, which can be more adaptive to the new anaerobic conditions,for example new leaves[7]or adventitious roots[25].However if the submergence is too long, the energy costs paid by biomass loss can not be compensated by the benefits of new tissue formation.If we consider the total amount of carbohydrates in a particular tissue (carbohydrate concentration × tissues dry mass), it will be evident that the carbohydrate storage in most tissues was greatly reduced in the submerged plants(data was not shown), since submergence caused a large reduction of carbohydrate concentration and more biomass loss.Therefore, reduced carbohydrate availability as well as much biomass loss could be the cause of mortality in the prolonged submergence treatment.Studies on other species showed that higher carbohydrate status was associated with higher flood-tolerance in tolerant sorghum varietyes[26]and some bottomland tolerant tree species[27].

Fig.3 Dry mass (g) in non-flooded (control), waterlogged (2 cm deep water above soil surface) and completely submerged (2 m deep water) S.variegata plants after 40, 90,120 and 180 days of treatments.Points =means ± SE, n = 7

Fig.4 Soluble sugar concentration and starch concentration (mg/g dry mass) in non-flooded(control), waterlogged (2 cm deep water above soil surface) and completely submerged (2 m deep water) S.variegata plants after 40, 90, 120 and 180 days of treatments.Points = means ± SE, n = 7
Generally, in the three Gorges Reservoir region, there were more flood-tolerant herbaceous species than woody plant species, probably due to the advantage of clonal growth for survival rate in the former group[28].Up to now,it has been shown thatArundinella anomala[1],Hemarthria altissima,Cynodon dactylon,Capillipedium assimile,Paspalum distichum[17,19]andPolygonum hydropiper[29]are all flood tolerant, while only three riparian shrubs was reported to be flood-tolerant:Myricaria laxiflora[18],Salix variegata[30]andDistylium chinense[31].However, in this study,S.variegataplants survived a little longer thanA.anomalaplants (Tab.1).Part of the reasons might be different carbohydrate distribution in the two species.S.variegatais a shrub species which stored high amount of carbohydrate in coarse roots and stems (Fig.4c—f); inA.anomalaplants, the carbohydrate storage was only in stems (Fig.3c &d), while in the roots the carbohydrate concentration was much lower compared with that ofS.variegataroots(Fig.2a & b, Fig.4a—d).Furthermore,after 180 days submergence, the soluble sugar or starch concentration in the roots ofA.anomalaplants was near to or quite below 10 mg/g dry mass (Fig.2a & b), while sugar or starch concentration was much higher in the fine roots ofS.variegata(Fig.4a&b, sugar:20 mg/g dry mass; starch: 100 mg/gdry mass).Under complete submer-gence conditions,carbon storage might be important to supply sugar for non-storage tissues for survival; when flooding recedes,new roots and leaves initiation for recovery growth also might depend on mobilization of carbohydrate storage.It was very likely that transportation of sugar from stems to roots were blocked in theA.anomalaplants due to oxygen deficiency[32].The cease of recharge of soluble sugar pool in the roots might finally lead to exhaustion of sugar and earlier death ofA.anomalaplants.

Tab.3 F-values and significance of a two-way ANOVA of soluble sugar concentration and starch in A.anomala and S.variegata plants in 3 water depth treatments: non-flooding (control), waterlogging (2 cm water above soil surface) and submergence (2 m deep water) after 40 days, 90 days, 120 days and 180 days of treatment
4 Conclusions
The results showed bothA.anomalaandS.variegataplants are very tolerant of long-term flooding.Both species are capable of mobilizing and utilizing carbohydrate storage up to at least 3 months of complete submergence.Coupling of biomass loss, reduction in carbohydrate concentration and mortality rate with increasing submergence duration suggest that high survival rate in these species might be closely associated with the limited but remarkable capacity to mobilize carbohydrate storage.
[1] Luo F L, Wang L, Zeng B,et al.Photosynthetic responses of the riparian plantArundinella anomalaSteud.in Three Gorges Reservoir region as affected by simulated flooding[J].Acta Ecologica Sinica, 2006, 26(11): 3602—3609 [羅芳麗, 王玲, 曾波, 等.三峽庫區(qū)岸生植物野古草(Arundinella anomalaSteud.)光合作用對水淹的響應(yīng).生態(tài)學(xué)報,2006, 26(11): 3602—3609]
[2] Jackson M B.Ethylene and responses of plants to soil waterlogging and submergence [J].Annual Review of Plant Physiology, 1985, (36): 145—174
[3] Cheng S P, Wu Z B, Xia Y C.Review on gas exchange and transportation in macrophytes [J].Acta Hydrobiologica Sinica, 2003, 27(4): 413—417 [成水平, 吳振斌, 琤夏宜.水生植物的氣體交換與輸導(dǎo)代謝.水生生物學(xué)報, 2003,27(4): 413—417]
[4] Bradford K J.Effects of soil flooding on leaf gas exchange of tomato plants [J].Plant Physiology, 1983, 73(2): 475—479
[5] Chen H J, Qualls R G, Blank R R.Effect of soil flooding on photosynthesis, carbohydrate partitioning and nutrient uptake in the invasive exoticLepidium latifolium[J].Aquatic Botany, 2005, (82): 250—268
[6] Vervuren P J A, Beurskens S M J H, Blom C W P M.Light acclimation, CO2response and long-term capacity of underwater photosynthesis in three terrestrial plant species [J].Plant Cell and Environment, 1999, 22(8): 959—968
[7] Mommer L, Visser E J W.Underwater photosynthesis in flooded terrestrial plants: a matter of leaf plasticity [J].Annals of Botany, 2005, 96(4): 581—589
[8] Gibbs J, Greenway H.Mechanisms of anoxia tolerance in plants.?.Growth, survival and anaerobic catabolism [J].Functional Plant Biology, 2003, 30(1): 1—47
[9] Greenway H, Gibbs J.Mechanisms of anoxia tolerance in plants.II.Energy requirements for maintenances and energy distribution to essential processes [J].Functional Plant Biology, 2004, 30(10): 999—1036
[10] Vartapetian B B, Jackson M B.Plant adaptation to anaerobic stress [J].Annals of Botany, 1997, 79(supplement A): 3–20
[11] Barclay A M, Crawford R M M.The effect of anaerobiosis on carbohydrate levels in storage tissues of wetland plants [J].Annals of Botany, 1983, 51(2): 255—259
[12] Schlüter U, Crawford R M M.Long-term anoxia tolerance in leaves ofAcorus calamusL.andIris pseudacorusL.[J].Journal of Experimental Botany, 2001, 52(364): 2213—2225[13] ?í?ková-Kon?alová H, Kv?t J, Thompson K.Carbon starvation: a key to reed decline in eutrophic lakes [J].Aquatic Botany, 1992, 43(2): 105—113
[14] Laan P, Blom C W P M.Growth and survival responses ofRumexspecies to flooded and submerged conditions:the importance of shoot elongation,underwater photosynthesis and reservecarbohydrates [J].Journal of Experimental Botany, 1990, 41(7): 775—783
[15] Ram P C, Singh B B, Singh A K,et al.Submergence tolerance in rainfed lowland rice: physiological basis and prospects for cultivar improvement through marker-aided breeding [J].Field Crops Research, 2002, 76(2—3): 131—152
[16] Das K K, Sarkar R K, Ismail A M.Elongation ability and non-structural carbohydrates levels in relation to submergence tolerance in rice [J].Plant Science, 2005, 168(1):131—136
[17] Wang H F, Zeng B, Li Y,et al.Effects of long term submergence on survival and recovery growth of four riparian plant species in Three Gorges Reservoir region, China [J].Chinese Journal of Plant Ecology, 2008, 32(5): 977—984 [王海鋒,曾波, 李婭, 等.長期完全水淹對 4種三峽庫區(qū)岸生植物存活及恢復(fù)生長的影響.植物生態(tài)學(xué)報, 2008, 32(5): 977—984]
[18] Chen F Q, Xie Z Q.Survival and growth responses ofMyricaria laxifloraseedlings to summer flooding [J].Aquatic Botany, 2009, 90(4): 333—338
[19] Tan S D, Zhang S J, Zhang K R,et al.Effect of long-time and deep submergence on recovery growth and photosynthesis of three grass species in Three Gorges Reservoir area [J].Journal of Wuhan Botanical Research, 2009, 27(4): 391—396 [譚淑端, 張守君, 張克榮, 等.長期深淹對三峽庫區(qū)三種草本植物的恢復(fù)生長及光合特性的影響.武漢植物學(xué)研究, 2009, 27(4): 391—396]
[20] Zhang Y H, Zeng B, Fu T F,et al.Effects of long-term flooding on non-structural carbohydrates content in roots ofSalix variegataFranch [J].Journal of Southwest China Normal University(Natural Science), 2006, 31(3): 153—156[張艷紅, 曾波, 付天飛, 等.長期水淹對秋華柳(SalixvariegataFranch) 根部非結(jié)構(gòu)性碳水化合物含量的影響.西南師范大學(xué)學(xué)報(自然科學(xué)版), 2006, 31(3): 153—156]
[21] Miller G L.Use of dinitrosalicylic acid regent for determination of reducing sugars [J].Analytic Chemistry, 1959, 31:426—428
[22] van Eck W H J M, Lenssen J P M, Rengelink R H J,et al.Water temperature instead of acclimation stage and oxygen concentration determines responses to winter floods [J].Aquatic Botany, 2005, 81(3): 253—264
[23] van Eck W H J M, van de Steeg H M, Blom C W P M,et al.Is tolerance to summer flooding correlated with distribution patterns in river floodplains? a comparative study of 20 terrestrial grassland species [J].Oikos, 2004, 107(2): 393—405
[24] Pierik R, van Aken J M, Voesenek L A C J.Is elongation-induced leaf emergence beneficial for submergedRumexspecies [J]?Annals of Botany, 103(2): 353—357
[25] Chen T, Zeng B, Ye, X Q,et al.Effect of flooding on adventitious root formation ofArundinella anomalaSteud.andSalix variegataFranch [J].Journal of Anhui Agricultural Science(China), 2007, 35(19): 5703—5704, 5712 [陳婷, 曾波, 葉小齊, 等.水淹對野古草和秋華柳不定根形成的影響.安徽農(nóng)業(yè)科學(xué), 2007, 35(19): 5703—5704, 5712]
[26] Singla N K, Jain V, Sawhney S K.Activities of glycoltic enzymes in leaves and roots of contrasting cultivars of sorghum during flooding [J].Biology Plantarum, 2003, 47(4):555—560
[27] Gravatt D A, Kirby C J.Patterns of photosynthesis and starch allocation in seedlings of four bottomland hardwood tree species subjected to flooding [J].Tree Physiology, 1998,18(6): 411—417
[28] Zhang X Y, Fan D Y, Xie Z Q,et al.Clonal integration enhances performance ofCynodon dactylonsubjected to submergence [J].Chinese Journal of Plant Ecology, 2010, 34(9):1075—1083 [張想英, 樊大勇, 謝宗強, 等.克隆整合有助于狗牙根抵御水淹.植物生態(tài)學(xué)報, 2010, 34(9): 1075—1083]
[29] Chen F Q, Li Y, Qie G W,et al.The morphological responses and endurance ofPolygonum hydropiperto flooding stress [J].Journal of Wuhan Botanical Research, 2008, 26(2): 142—146[陳芳清, 李永, 郄光武, 等.水寥對水淹脅迫的耐受能力和形態(tài)學(xué)響應(yīng).武漢植物學(xué)研究, 2008, 26(2): 142—146]
[30] Li Y, Zeng B, Ye X Q,et al.The effects of flooding on survival and recovery growth of the riparian plantSalix variegataFranch.in Three Gorges reservoir region [J].Acta Ecologica Sinica, 2008, 28(5): 1923—1930 [李婭, 曾波, 葉小齊, 等.水淹對三峽庫區(qū)岸生植物秋華柳(Salix variegataFranch.)存活和恢復(fù)生長的影響.生態(tài)學(xué)報, 2008,28(5): 1923—1930]
[31] Chen G F, Cai K Y, Li Z J,et al.Flooding effect on growth and physiology ofDistylium chinesis[J].Journal of Southwest Forestry College, 2008, 28(5): 42—44 [陳桂芳, 蔡孔瑜, 李在軍, 等.淹水對中華蚊母樹生長及生理的影響.西南林學(xué)院學(xué)報, 2008, 28(5): 42—44]
[32] van Dongen J T, Schurr U, Pfister M,et al.Phloem metabolism and function have to cope with low internal oxygen [J].Plant Physiology, 2003, 131(4): 1529—1543
猜你喜歡
汽車實用技術(shù)(2022年16期)2022-09-03 01:28:09
四川地質(zhì)學(xué)報(2020年2期)2020-05-31 06:53:00
小哥白尼(軍事科學(xué))(2018年3期)2018-06-15 09:56:56
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
山西農(nóng)經(jīng)(2016年3期)2016-02-28 14:23:53
少兒科學(xué)周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學(xué)周刊·兒童版(2015年1期)2015-07-07 04:12:52
河南科技(2015年3期)2015-02-27 14:20:52
天然氣勘探與開發(fā)(2012年2期)2012-03-25 12:56:43
