
The role of microRNAs during the genesis of medulloblastomas induced by the hedgehog pathway
2011-11-02 07:33:18XiaojuLuoJunLiuStevenCheng
Xiaoju Luo, Jun Liu, Steven Y Cheng
Department of Developmental Genetics and Center for Regenerative Medicine, Nanjing Medical University, Nanjing, Jiangsu 210029, China
INTRODUCTION
Hedgehog (Hh)is one ofthe primary morphogenic signals that specify the patterns of cell growth and differentiation during vertebrate development. Sustained Hh signaling pathway activity can lead to congenital anomalies and development of various tumors[1].Medulloblastomas (MB), which are aggressive childhood brain tumors of cerebellar origin, are associated with inappropriate Hh pathway activity[2-5]. MB is a"stem cell" tumor caused by the abnormal proliferation of granular cells in the internal granule layer (IGL)of the cerebellum, which come from the differentiation of granular cell precursors (GCPs)in the external germinal layer (EGL)[6-7]. In general, the GCPs stop dividing and exit the cell cycle to keep the number of granular cells at a stable level one month after birth.When the Hh pathway is abnormally activated, GCP cells cannot exit the cell cycle; however, they continue to proliferate and form a large number of granule cells accumulating in the EGL layer, and finally develop MB. The mechanism of this pathological process remains elusive[8-9].
MicroRNAs (miRNAs)are a group of small noncoding RNAs that are about 22-nucleotide in length and can silence gene expression by mRNA degradation or translation inhibition through binding to the 3’untranslated region (3’UTR)of target mRNAs[10-12].The relationship between the Hh signaling pathway and miRNAs has been reported. Flynt et al.[13]discovered that miR-214 targeted a Hh pathway suppressor gene Sufu to regulate somite development in zebrafish.Lee et al.[14]reported that miRNA-378 promoted cell survival, tumor growth and angiogenesis by targeting SuFu. Paul et al.[15]showed that the miRNA17-92 cluster collaborated with the Hh pathway in medulloblastoma. In the present study, we chose 90 miRNA candidates to screen out the miRNAs probably regulated by the Hh signaling pathway and to further clarify the role of miRNAs in the genesis of MB induced by abnormal Hh signaling pathway activity.
MATERIALS AND METHODS
Cell culture
Wildtype (WT)mouse embryonic fibroblast (MEF),Ptc-/- MEF and Sufu-/- MEF cells were maintained in DMEM containing 100 U/mL penicillin-streptomycin supplemented with 10% fetal bovine serum(FBS, Gibco, USA)at 37°C in 100-mm cell culture dishes (Corning, USA)in a humidified atmosphere of 5% CO2. When cells reached confluence, they were washed with 0.01 mol/L PBS, digested with 0.25%trypsin-EDTA for 2 min, centrifuged at 1,500 g for 3 min, and then resuspended in culture medium.
Animals
Ptc and p53 gene knockout C57/B6 mice (gifts from the Jackson laboratory)were housed in a temperatureand humidity-controlled room and maintained on a 12-h light-dark cycle with access to food and water ad libitum. These two kinds of knockout mice (Ptc and p53 gene knockout mice)were mated to form hybrids,respectively.
Design of RT-PCR primers
Mature miRNAs were amplified by the method of stem-loop RT-PCR, which was firstly invented and reported by Chen et al.[16]Through adding an additional stem-loop structure at the 5’ end of the reverse transcription primers, it can increase the length of cDNA and steric hindrance to increase the specificity of PCR amplification of short RNA fragments. The PCR primers of Gli1 and GAPDH were designed using the Primer 5.0 software. The primer sequences were: Gli1:5’-TCCAGCTTGGATGAAGGACCTTGT-3’ (sense)and 5’-GCATATCTGGCACGGAGCATGTA-3’ (antisense); GAPDH: 5’-ACCCAGTCCTCACCTTCCAC-3’(sense)and 5’- GGCCTCCTCTTTCTCCCAC-3’ (antisense).
RNA isolation and RT-PCR
Total RNAs were extracted from MEF cells and mouse normal and tumor tissues with RNAiso Plus reagent (Takara, Japan)according to the manufacturer’s instructions. A 0.5 μg aliquot of total RNAs was reverse transcribed into cDNA using the stem-loop reverse transcript primers mentioned before. Then,mature miRNAs were amplified by PCR using the appropriate primers. The PCR was carried out in a total volume of 20 μL containing 20 mmol/L Tris-HCl, 50 mmol/L KCl, 1.5 mmol/L MgCl2, 0.2 mmol/L dNTPs,0.6 mmol/L of forward and reverse primers, and 2.5 U of Taq DNA polymerase. The housekeeping gene-GAPDH was used as an internal control. MiR-10a and miR-125b were the internal controls of miRNAs. PCR was done in PCR tube, and the amplification cycles of Gli1 and GAPDH were 35 and 26, respectively.Denaturing, annealing, and extension reactions were performed at 94°C for 30 s, 55°C for 30 s, and 72°C for 45 s, respectively. Amplification condition of miRNAs was 95°C for 5 min followed by 40 cycles of 95°C for 15 s and 60°C for 1 min.
Mouse breeding and extraction of medulloblastomas
Mice homozygous for Ptc mutation died before E9.5 because of closed neural tube defects. We got Ptc+/- mice through mating male Ptc+/- and female Ptc+/- mice. Ptc+/- mice were also crossed with p53-/-mice to get p53+/-; Ptc+/- or p53-/-; Ptc+/- mice. The status of mice was carefully observed during the experiment process. As soon as the anomalous cerebellar symptoms such as skull enlargement, slow response or ataxia appeared, the mice were sacrified and the tumor tissues were removed for total RNAs isolation. Compared to the normal cerebellums, the tumor tissues showed increased volume and flattened superficial sulcus.
The animal protocal was approved by the local institation review board and animal study was carried out in accordance with the established guidelines for experimental animal use.
Statistical analysis
Values were shown as mean±SD. The primary screening was performed by one-way ANOVA and Bonferroni adjustment. P value < 0.017 was considered statistically significant. Student’s t-test was used to test the expression of the miR-154 cluster. P value< 0.05 was considered statistically significant. All the statistical analyses were performed with SPSS11.0(SPSS InC., USA).
RESULTS
Screening of miRNAs regulated by the Hh signaling pathway in WT MEF, Ptc-/-MEF,and Sufu-/-MEF cell lines
Ptc and Sufu are two important suppressors of the Hh signaling pathway. When one of the two genes is knocked out, the Hh signaling pathway will be activated. Gli1, the downstream transcription factor of Hh pathway, can be considered as the marker of the Hh pathway activity[17-21]. At first, we detected the Gli1 expression in WT MEF, Ptc-/- MEF and Sufu-/- MEF cells, and found that the Gli1 mRNA levels were higher in the two knockout cell lines; in other words, the Hh pathway was activated in them (Fig. 1A). Then, we tested the expression of 90 different miRNAs in WT MEF and Ptc-/- MEF cell lines by the stem-loop RTPCR. The results showed that 15 miRNAs showed differential expression in the two MEF cell lines (Fig.1B). But only 13 miRNAs had significantly differential expression, they were let-7e, miR-30b, miR-30c,miR-34a, miR-96, miR-129-3p, miR-132, miR-134,miR-135a, miR-143, miR-146a, mkiR-154 and miR-183, respectively (Fig. 1C). Afterwards, we performed secondary screening in Sufu-/- MEF cells through the stem-loop RT-PCR. The 15 miRNAs from primary screening were tested and the result was quantified with ImageJ. Of the miRNAs examined, 10 had the same expression trend and significant difference in the two knockout cell lines, and they were let-7e, miR-30b, miR-30c, miR-96, miR-129-3p, miR-132, miR-134, miR-135a, miR-143 and miR-183, respectively(Fig. 1C).
Quantification of expression of miRNAs for validation of screening results in mouse MB and normal cerebellar tissues
It is well known that Ptc is an important suppressor of the Hh signaling pathway. MB is extremely frequent in Ptc knockout mice. Ptc-/- mice died at E9.5, while Ptc+/- mice were viable and susceptible to MB[22-23]. In the current study, 20%-30% Ptc heterozygous mice developed brain tumors at the age of about 5 weeks. Then, we separated Ptc+/- MB and normal cerebellar tissues from B6/C57 mice (Fig. 2A and Fig. 2B). Total RNAs were isolated from these tissues and the mRNA levels of Gli1 were detected by RT-PCR. The Gli1 mRNA levels of the Ptc+/- MB were higher than those of the normal cerebellums (data not shown). Therefore, it suggested that Hh signaling was overactivated in the tumor tissues. Then, we validated the expression of the 15 miRNAs selected by the primary and secondary screening. The results showed that miR-134 and miR-154 were down-regulated in the MB, but the other miRNAs showed no significant difference in expression (Fig. 2C). In order to verify the above result, we examined the expression of miR-134 and miR-154 in the other four groups of tumor and normal tissues. MiR-154 was down-regulated in the other four MB, but the change of miR-134 was insignificant (Fig. 2D).
p53 is a tumor suppressor protein. The frequency of MB in Ptc+/- mice was increased by a p53 mutant background[24]. Indeed, there were about 70%-80% incidence rate of MB in the p53 and Ptc double knockout mice. In order to further validate the results in the Ptc+/- mice, we repeated the experiment in MB with p53+/- Ptc+/- and p53-/- Ptc+/-. The Hh signaling pathway was activated in p53 and Ptc double knockout MB with a dramtically decrease of miR-154 levels(Fig. 2E). This result indicated that the expression of miR-154 was also significantly down-regulated in the MB which were induced by activated Hh pathway activity. MiR-154 may play an important role in the formation of MB induced by activated Hh signaling.
Expression profile of themiR-154cluster during the genesis of MB
Mouse genomic structure analysis showed that miR-134 and miR-154 were located in the same miRNA cluster on chromosome 12. The miR-154 cluster is a very huge and complicated gene cluster, including more than 40 miRNA members. To make clear whether other members of the miR-154 cluster had the same expression profile in MB, we detected the other 21 members of the miR-154 cluster by the stem-loop RT-PCR. Seven of them were undetectable because of low expression levels (data not shown). Of the 14 miRNAs, 6 were down-regulated, and 1 was up-regulated in the MB tissues (Fig. 3). The result indicated that not only miR-154 but also the whole cluster was regulated by the Hh signaling pathway, and miR-154 may play an important role during the genesis of MB.
DISCUSSION
MiRNAs play a critical role in developmental and physiologic processes and are involved in the pathogenesis of several human diseases including cancer[25-27].Recently, the relationship between the Hh signaling pathway and miRNAs has been reported[13-15]. Search of miRNAs that are regulated by the Hh pathway can help us better understand the Hh pathway, clarify the molecular mechanism of MB, and finally explore new therapeutic targets to cure diseases associated with the Hh pathway.
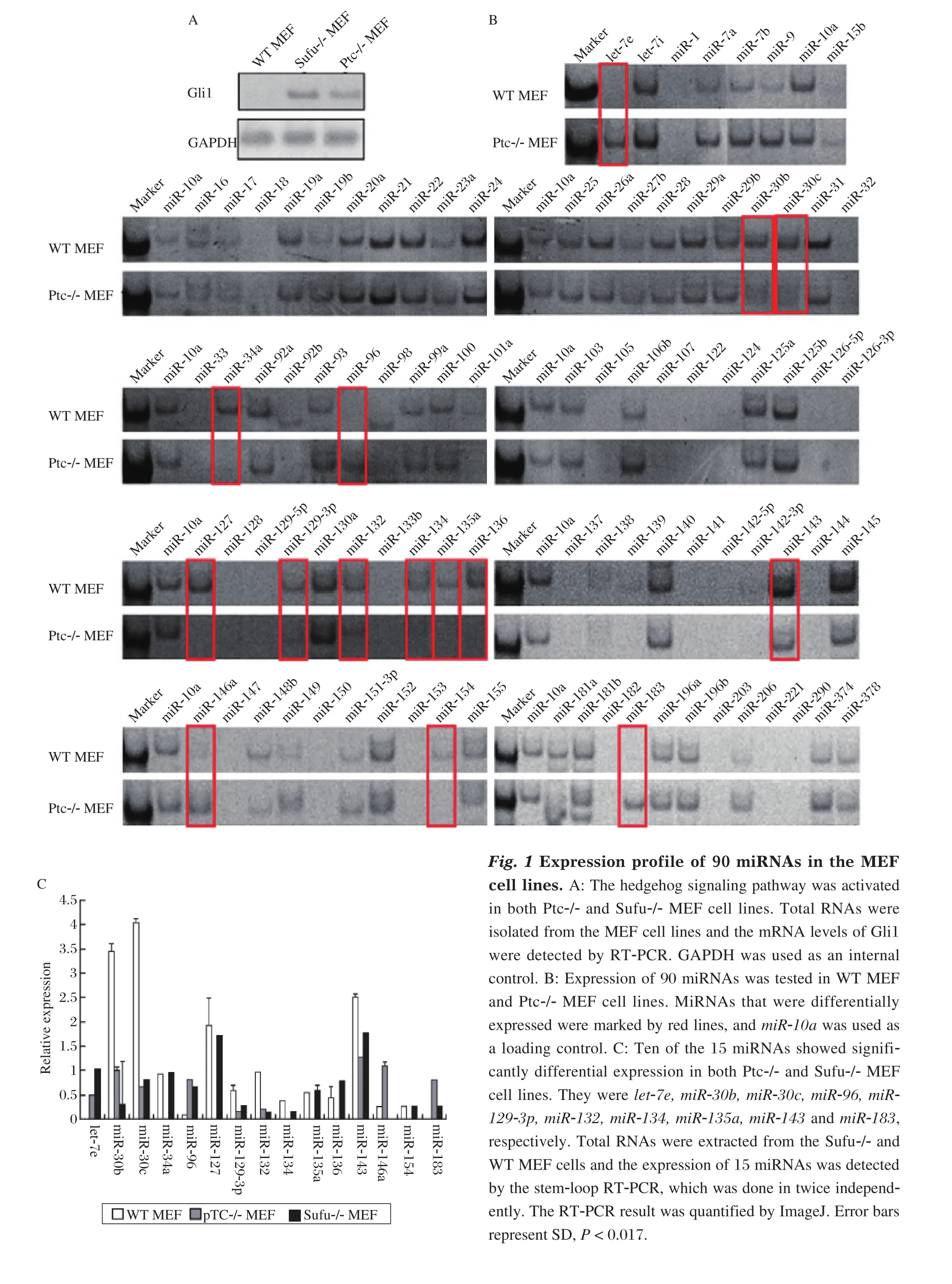
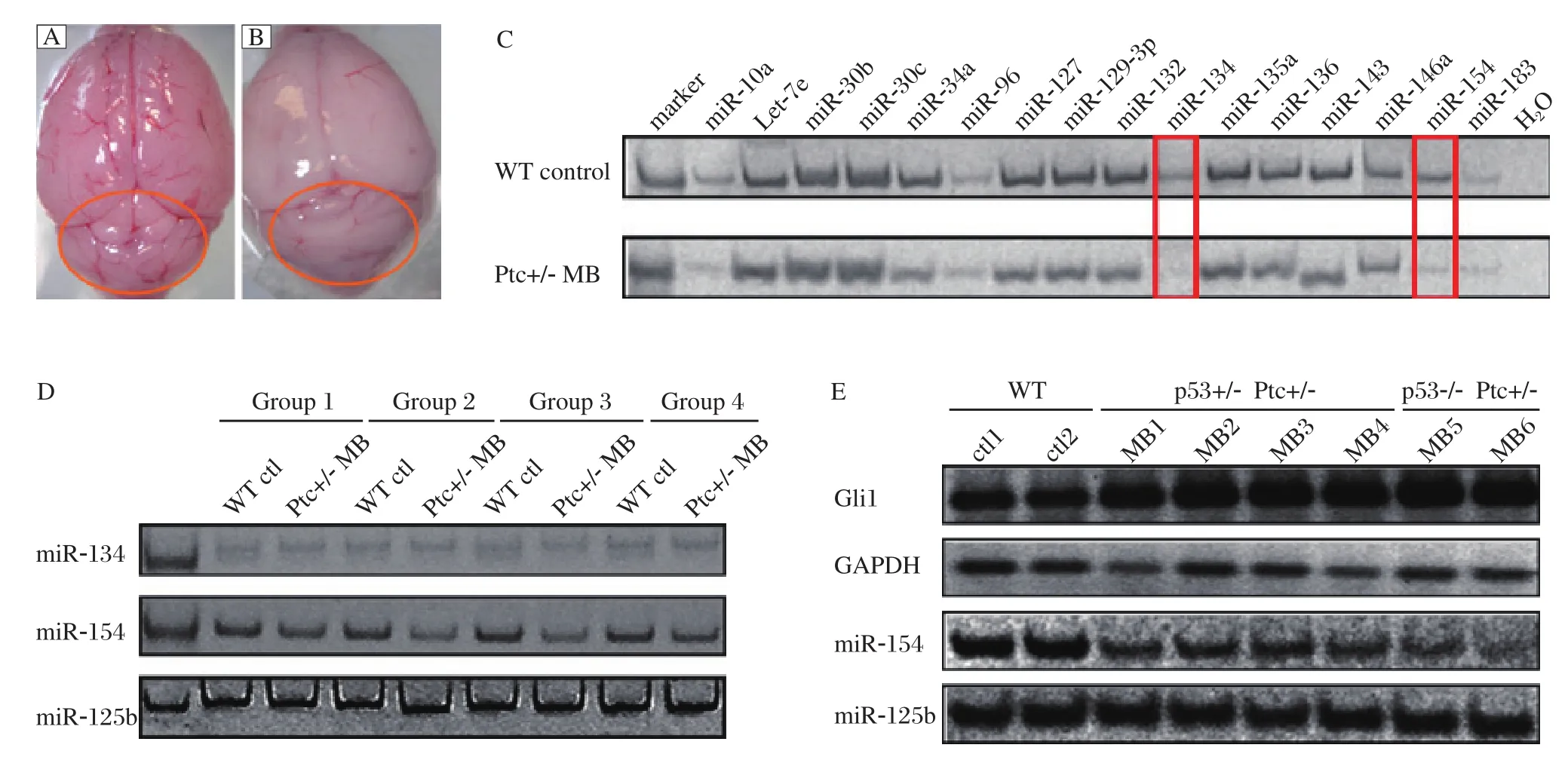
Fig. 2 Comparison between normal and medulloblastomas (MB)samples revealed changes in expression of miRNAs. Normal cerebellar tissue (A)and MB (B)are shown here. The areas marked by the red lines indicate the cerebellar tissues, where MB usually develop. C: Fifteen miRNAs identified from the primary screening were tested by stem-loop RT-PCR in the MB and normal cerebellar tissues with miR-10a as an internal loading control and H2O as the negative control. D: Validation of the expression of miR-134 and miR-154 in the other four groups of Ptc+/- MB and WT normal tissues. MiR-125b was used as a loading control. E: Expression of miR-154 in p53+/- Ptc+/- and p53-/- Ptc+/- MB. Gli1 expression was also detected. MiR-125b was used as a loading control. WT: wildtype; ctl: control.
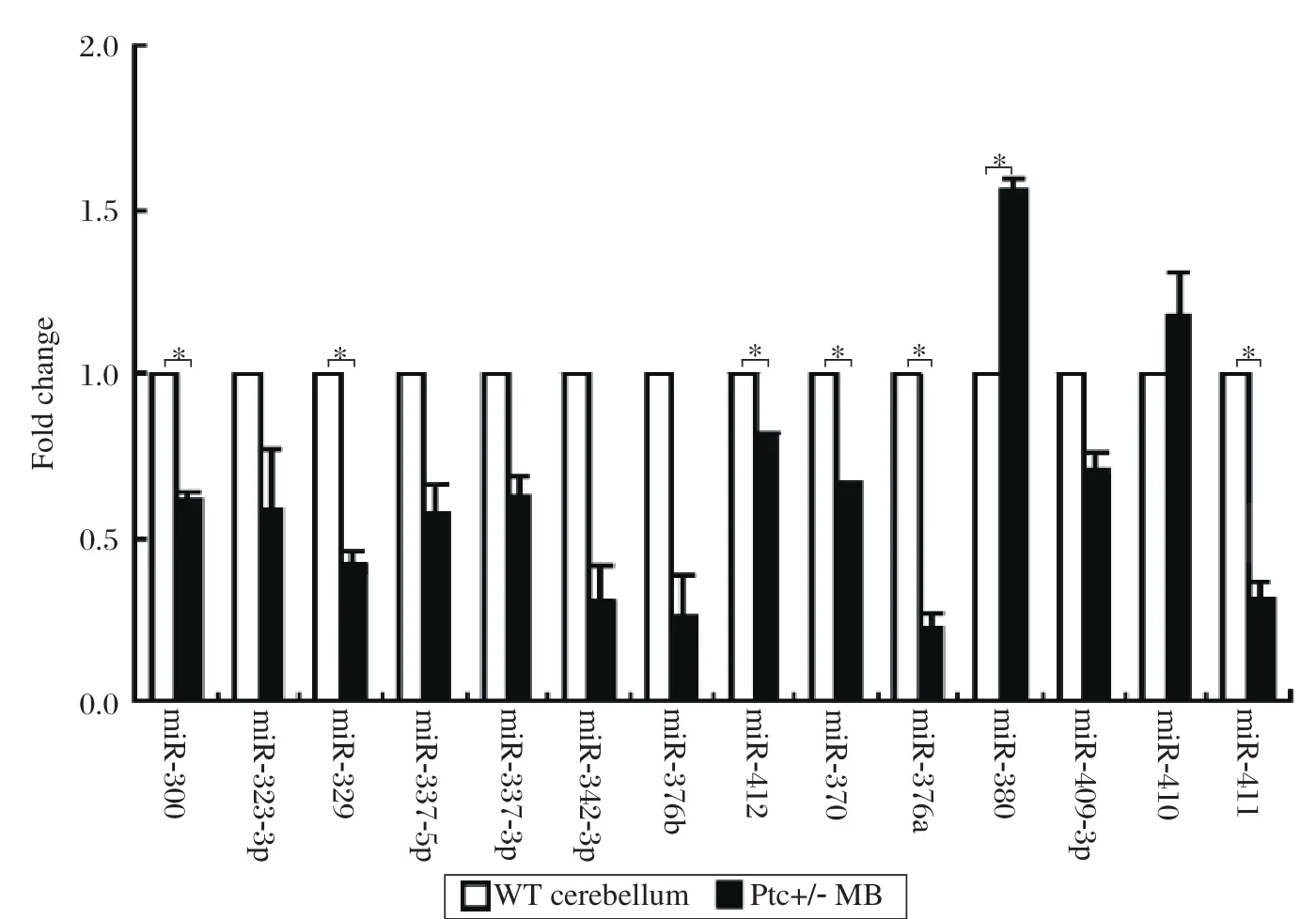
Fig. 3 Relative expression of 14 members of the miR-154 cluster in Ptc+/- medulloblastomas (MB)and wildtype(WT)normal cerebellar tissues. The RT-PCR result was quantified by ImageJ software. Six of them were down-regulated,while 1 of them was up-regulated in the MB. This experiment was done twice independently. Error bars represent SD, *P < 0.05.
We selected different MEF cell lines to carry out screening because they are sensitive to the Hh signaling pathway. In the current study, we found that 13 miRNAs showed significantly differential expression between the WT MEF and Ptc-/- MEF cell lines.Then, we performed secondary screening in the Sufu-/- MEF cell lines and the result showed that 10 of the 15 miRNAs had the same expression trend both in Ptc-/- MEF and Sufu-/- MEF cell lines. These 10 miRNAs are interesting because they probably are regulated by the Hh pathway. However, it is still unknown whether they are directly regulated by the Hh pathway or not.This needs more studies on gene transcription regulation. The expressly of the other 5 miRNAs were inconsistent in the Ptc-/- and Sufu-/- MEF cell lines, and it may be falsely positive in the screening.
Subsequently, we verified the 15 miRNA candidates in mouse MB which were induced by activated Hh signaling and found that the expression of miR-154 was significantly lower in the tumor samples than in the normal tissues, in spite of the genotypes. Our study indicated that miR-154 was closely related to the Hh signaling pathway, and it played an essential role in the genesis of MB induced by activated Hh signaling. There are many miRNAs in the miR-154 cluster,but the other members may not be involved in this process.
MB is the most common brain tumor in children,accounting for 10%-20% of the central nervous system tumors. According to the statistics, the peak age of onset of MB is 7 years. Clinically, MB is a highly malignant tumor with aggressive biological behavior and high mortality rate[28]. To improve the survival rate, investigators have carried out numerous studies at the molecular level to explore the pathogenesis of this tumor. Many studies show that the vast majority of MB are induced by sustained activation of the Hh pathway, but the molecular mechanism of MB occurrence has not been characterized yet. We tried to do some research on this problem from the angle of miRNAs.
The miR-154 cluster, located in the human imprinted 14q32 domain (mouse chromosome 12F2), is a very conservative miRNA cluster in mammalians[29].It is also a very complex and large cluster, including more than 40 members spanning over a distance of over 40 kb. Because of its complex structure, the current study of the miR-154 cluster is relatively limited.The transcription regulation of this cluster is poorly known. It has been considered to play a role in the epigenetic modifications and regulation of embryonic development[30,31]. In this study, we reported that the miR-154 cluster played a role in medulloblastoma genesis, but it requires further studies on the regulation mechanism of miR-154 by the Hh signaling pathway and further validation of our screening results in more types of tumor.
Reference
[1]Ferretti E, Smaele ED, Miele E, Laneve P, Po A, Pelloni M. Concerted microRNA control of Hedgehog signalling in cerebellar neuronal progenitor and tumour cells.Embo J 2008;27:2616-27.
[2]Goodrich LV, Milenkovic L, Higgins KM, Scott MP. Altered neural cell fates and medulloblastoma in mouse patched mutants. Science 1997;277:1109.
[3]Reifenberger J, Wolter M, Weber RG, Megahed M, Ruzicka T, Lichter P, et al. Missense Mutations in SMOH in Sporadic basal cell carcinomas of the skin and primitive neuroectodermal tumors of the central nervous system. Cancer Res 1998;58:1798.
[4]Wolter M, Reifenberger J, Sommer C, Ruzicka T, Reifenberger G. Mutations in the human homologue of the drosophila segment Polarity gene patched (PTCH)in sporadic basal cell carcinomas of the skin and primitive neuroectodermal tumors of the central nervous system.Cancer Res 1997;57:2581.
[5]Xie JW, Johnson RL, Zhang XL, Bare JW, Waldman FM,Cogen PH, et al. Mutations of the PATCHED gene in several types of sporadic extracutaneous tumors. Cancer Res 1997;57:2369.
[6]Barisone GA, Yun JS, Diaz E. From cerebellar proliferation to tumorigenesis: new insights into the role of Mad3. Cell Cycle 2008;7:423-7.
[7]Wechsler Reya RJ, Scott MP. Control of neuronal precursor proliferation in the cerebellum by Sonic Hedgehog.Neuron 1999;22:103-14.
[8]Oliver TG, Read TA, Kessler JD, Mehmeti A, Wells JF, Huynh TT, et al. Loss of patched and disruption of granule cell development in a pre-neoplastic stage of medulloblastoma. Development 2005;132:2425-39.
[9]Cheng SY, Yue S. Role and regulation of human tumor suppressor SUFU in Hedgehog signaling. Adv Cancer Res 2008;101:29-43.
[10]Kiriakidou M, Tan GS, Lamprinaki S, De Planell-Saguer M, Nelson PT, Mourelatos Z. An mRNA m7G cap binding-like motif within human Ago2 represses translation.Cell 2007;129:1141-51.
[11]Bagga S, Bracht J, Hunter S, Massirer K, Holtz J, Eachus R, et al. Regulation by let-7 and lin-4 miRNAs results in target mRNA degradation. Cell 2005;122:553-63.
[12]Humphreys DT, Westman BJ, Martin DI, Preiss T. MicroRNAs control translation initiation by inhibiting eukaryotic initiation factor 4E/cap and poly(A)tail function. Proc Natl Acad Sci 2005;102:16961-6.
[13]Flynt AS, Li N, Thatcher EJ, Solnica-Krezel L, Patton JG. Zebra fi sh miR-214 modulates Hedgehog signaling to specify muscle cell fate. Nature genetics 2007;39(2):259-63.
[14]Lee DY, Deng Z, Wang CH, Yang BB. MicroRNA-378 promotes cell survival, tumor growth, and angiogenesis by targeting SuFu and Fus-1 expression. Proc Natl Acad Sci 2007;104:2350-5.
[15]Northcott PA, Fernandez A, Hagan JP, Ellison DW,Grajkowska W, Gillespie Y, et al. The miR-17/92 Polycistron is up-regulated in Sonic Hedgehog-driven medulloblastomas and induced by N-myc in Sonic Hedgehog-treated cerebellar neural precursors. Cancer Res 2009;69:3249-55.
[16]Chen C, Ridzon DA, Broomer AJ, Zhou Z, Lee DH,Nguyen JT, et al. Real-time quantification of microRNAs by Stem-loop RT-PCR. Nucleic Acids Res 2005;33:179.
[17]Pazzaglia S. Ptc1 heterozygous knockout mice as a model of multi-organ tumorigenesis. Cancer Lett 2006;234:124-34.
[18]Raffel C, Jenkins RB, Frederick L, Hebrink D, Alderete B, Fults DW, et al. Sporadic medulloblastomas contain PTCH mutations. Cancer Res 1997;57:842-5.
[19]Vorechovsky I, Tingby O, Hartman M, Stromberg B,Nister M, Collins VP, et al. Somatic mutations in the human homologue of Drosophila patched in primitive neuroectodermal tumours. Oncogene 1997;15:361-6.
[20]Taylor MD, Liu L, Raffel C, Hui CC, Mainprize TG,Zhang X, et al. Mutations in SUFU predispose to medulloblastoma. Nat Genet 2002;31:306-10.
[21]Cheng SY, Yue S. Role and regulation of human tumor suppressor SUFU in Hedgehog signaling. Adv Cancer Res 2008;101:29-43.
[22]Goodrich LV, Milenkovic L, Higgins KM, Scott MP.Altered neural cell fates and medulloblastoma in mouse patched mutants. Science 1997;277:1109-13.
[23]Hahn H, Wojnowski L, Zimmer AM, Hall J, Miller G,Zimmer A. Rhabdomyosarcomas and radiation hypersensitivity in a mouse model of Gorlin syndrome. Nature Med 1998;4:619-22.
[24]Wetmore C, Eberhart DE, Curran T. Loss of p53 but not ARF accelerates medulloblastoma in mice heterozygous for patched. Curran, Cancer Res 2001;61:513.
[25]Gaur A, Jewell DA, Liang Y, Ridzon D, Moore JH, Chen CF, et al. Characterization of MicroRNA Expression Levels and Their Biological Correlates in Human Cancer Cell Lines. Cancer Res 2007;67:2456.
[26]Lu J, Getz G, Miska EA, Alvarez-Saavedra E, Lamb J,Peck D, et al. MicroRNA expression pro fi les classify human cancers. Nature 2005;435:834-8.
[27]Espinosa CE, Slack FJ. The role of microRNAs in cancer. Yale J Biol Med 2006;79:131-40.
[28]Reddy AT, Packer RJ. Pediatric central nervous system tumors. Curr Opin Oncol 1998;10:186-93.
[29]Lin SP, Youngson N, Takada S, Seitz H, Reik W, Paulsen M, et al. Asymmetric regulation of imprinting on the maternal and paternal chromosomes at the Dlk1-Gtl2 imprinted cluster on mouse chromosome 12. Nat. Genet 2003;35:97-102.
[30]Mourelatos Z, Dostie J, Paushkin S, Sharma A, Charroux B, Abel L, et al. miRNPs: A novel class of ribonucleoproteins containing numerous miRNAs. Genes&Dev 2002;16:720-8.
[31]Williams AE, Moschos SA, Perry MM, Barnes PJ, Lindsay MA. Maternally imprinted miRNAs are differentially expressed during mouse and human lung development.Dev& Dyn 2006;236:572-80.
 THE JOURNAL OF BIOMEDICAL RESEARCH2011年1期
THE JOURNAL OF BIOMEDICAL RESEARCH2011年1期
- THE JOURNAL OF BIOMEDICAL RESEARCH的其它文章
- Development and optimization of an antibody array method for potential cancer biomarker detection☆
- Single-dose and multiple-dose pharmacokinetics of zaltoprofen after oral administration in healthy Chinese volunteers
- A mutation in the type II hair keratin KRT86 gene in a Han family with monilethrix☆
- Instructions for Authors
- 《生物醫(yī)學(xué)研究雜志(Journal of Biomedical Research)》宣布與湯森路透合作——采用國(guó)際一流的在線投審稿系統(tǒng)ScholarOne Manuscripts
- Successful conservative management with methotrexate and mifepristone of cervical pregnancy